
Kiwifruit is not covered in hairs. It’s covered in trichomes. And only if you’re talking about green Actinidia chinensis var. deliciosa. But, why? One answer is: pretty much to keep it from drying out. Another is: because it’s a polyploid from western China and was kind of chosen at random to be the most commonly grown kiwifruit, and they’re not all fuzzy. Those aren’t mutually exclusive answers. Put on your ecophysiology hats and grab a paring knife.
Think of fruit growth as a balancing act between ingoing and outgoing fluxes. When the balance is positive, fruits grow. When it is negative, they shrink—or shrivel. The main fluxes in question are carbon and water, which enter the fruit from the xylem and phloem of the plant vascular system. Water is lost mainly to the atmosphere via transpiration (evaporative water lost through stomata and other pores and from the skin surface). Keeping the ledger positive isn’t an easy job for a fruit. Hot, dry, and windy weather encourages transpiration and thereby increases the odds that a fruit will experience water stress. Excessive sunlight may cause sunburn. Fruits also need to avoid attack from pathogens and herbivores before the seeds within mature. A fruit’s skin—its cuticle and epidermis—is its first line of defense against abiotic and biotic threats. Some fruits resort to creative coverings to get the job done.
Here I’ll take a close look at the skin of kiwifruits. Why, exactly, are they so fuzzy?

A heart-shaped green kiwifruit (Actinidia chinensis var. deliciosa), covered in fuzzy trichomes
Functional fuzz
The short explanation for why kiwifruits are fuzzy is that they are covered with trichomes: hair-like extensions arising from the cell walls of the epidermis whose structure can vary widely. The trichomes covering kiwifruit are multicellular, and generally come in short and long varieties (1). Kiwifruit’s trichomes are also nonglandular, as opposed to specialized glandular trichomes that bear essential oils (see examples in our essays on lemon flavor and green walnut), or stinging trichomes, like those on nettle (Urtica spp.).
The fuzziest kiwifruit most of us will encounter is the widely commercially available green kiwifruit, cultivars of Actinidia chinensis var. deliciosa, which has bright green flesh when ripe (we’ll explore green fruit color in an upcoming post) and tough, inedible skin covered with coarse fuzz (trichomes). The length and density of trichomes actually varies within and across the 50+ species in the kiwifruit genus Actinidia (family Actinidiaceae, order Ericales; 2–4). For example, yellow-fleshed “golden” kiwifruit (A. chinensis var. chinensis), which is becoming increasingly commercially available, is covered less densely and with shorter trichomes than green kiwifruit (1). And if you’re especially lucky you might get your hands on tiny kiwiberries (Actinidia arguta), which have a hairless, thin, edible skin, much like a grape. This trichome variation is part of the longer explanation of why (some, most) kiwifruits are densely covered with trichomes, which is that trichomes help defend the fruit against adverse environmental conditions.

Mature fruits from (A) A. chinensis var. deliciosa (fruit color green), (B) A. chinensis var. chinensis (golden kiwi, fruit color yellow), and (C) A. arguta (kiwiberry). Bar = 10mm. Notice the prominent persistent stigmas on the distal end (opposite the stem) and persistent calyces at the stem end. From Hallet and Sutherland (2005).
Just like any other would-be herbivore, you likely experience kiwifruit trichomes as an annoying impediment in your quest to access the fruit within. As to that fruit within, the botany (anatomy, breeding system) of kiwifruit is a bit tangential to this discussion of its trichome ecology (its relationship to the environment and other organisms), but as a hopefully useful aside, you can find the botany basics of kiwifruit in Figures 1 and 2 below.

Figure 1. Position of major vascular bundles (xylem and phloem) and structures in kiwifruit. Kiwifruit is botanically classified as a berry, a fleshy fruit that develops from a single ovary. The kiwifruit ovary is composed of numerous fused carpels. Each carpel bears its own style and stigma, creating a starburst pattern in the center of the pistillate flowers (see Figure 2 below). Those stigmas can persist through fruit ripening and remain visible at an end of ripe kiwifruit (shown here on A. arguta). The same goes for the tough sepals below the petals. Those many carpels contain chambers called locules, in which seeds develop. Septa divide locules and carpels within the ovary and are visible in a kiwi cut in cross section as pale white lines (shown here for A. chinensis var. deliciosa). The carpels radiate around the large, white central (axial) placenta, which is formed of homogeneous, large parenchyma cells and vascular tissue. The major vascular bundles in the fruit are also generally visible in cross section as translucent dots. Anatomy from Clearwater et al. (2012) and Guo et al. (2013).

Figure 2. All species in the kiwifruit genus (Actinidia) are climbing woody vines (lianas) that are dioecious, with separate staminate (male) and pistillate (female) flowers, usually borne on separate vines. The pistillate Actinidia flower (photo from Guo et al. 2013) in the photos shows multiple stigmas and styles arising from the fused carpels in the ovary (the pistillate flowers do make stamens but lack viable pollen). Each stigma/style provides an avenue for the pollination of the ovules in the locules of one of the fused carpels within the ovary. The stigmas are persistent and visible on ripe fruit.
Bugs, weather, or both
The nonglandular trichomes covering kiwifruits are similar to those creating fuzz on sundry other structures in various species: think of fuzzy leaves, stems, bracts, buds, inflorescences you may have seen. Despite the ease with which examples of fuzzy plant structures come to mind, it is not always straightforward to infer the function of that fuzz. Most of the function of glandular trichomes appears to be defense against herbivores and pathogens, mostly by way of the noxious chemicals that the glandular trichomes produce. By simply creating a physical barrier, defense might also account for some or all of the purpose of nonglandular trichomes. There is some evidence for that, at least for fuzzy leaves in some species (5–7). After incurring herbivore damage, for example, several species have been observed to increase the density or number of trichomes covering new leaves (8, 9). The primary function of fuzz-forming trichomes, though, may also be to help plants cope with stress from prevailing climatic conditions, and that is likely the case for the fuzzy fruits in question here. Keep in mind, though, that trichomes may serve more than one purpose, and I don’t know of any studies for any trichome-covered structure in which hypotheses regarding pests and weather have been tested simultaneously.

Persistent sepals on a green kiwifruit
Fuzzy logic
Fuzzy leaves and other structures are more well-studied than fuzzy fruits, but most evidence available points to broadly similar function of dense trichomes on both structures. The problem is, you can find trichome-covered plant structures in almost every environment on Earth where plants grow, and it seems that the primary function of the fuzziness varies across those many environments because the dangers to plants vary, too. Not all of the plants in a given spot will have obviously fuzzy parts, though, which is a clue that trichome density is only one plant characteristic that a plant species may have modified in order to live under the prevailing conditions. Trichome density, at least on leaves, too, is plastic: it varies in response to recent environmental conditions, including weather and herbivory, not just overall climate trends. That is, in a dry year, some plant species may increase trichome density, and we’ve already described how plant structures re-growing after herbivory may have higher trichome density.

Alpine Pedicularis lanata and Dryas, with woolly trichome-covered inflorescence and leaves, respectively (Chugach Mountains, Alaska)
It is likely that trichome density is idiosyncratic to individual plant populations (10), species, or lineages (11–13), and much work remains to piece together its evolutionary trends. In general, trichomes appear to moderate the ravages of various environmental stresses by affecting the temperature and humidity of air in the boundary layer at the plant surface (9, 14–16), or by reflecting excessive sunlight or ultraviolet radiation (17–21).
Some trichomes might act as umbrellas, repelling excess water (and perhaps fungal spores) from plant surfaces in overly wet environments (22), while in dry or foggy environments, trichomes might help with water uptake (23, 24) or humidity maintenance (25–30). In hot environments, trichomes may help cool the plant surface by reflecting sunlight, reducing the plant’s need to dissipate excess heat via evaporative cooling, and thereby reducing water loss (21). In cold environments, fuzzy trichomes act in part as an insulative blanket, warming the underlying structures. Think of the fuzzy bracts covering flower buds in magnolia and pussywillows (22, 31) emerging in the cool early spring.
From leaves to peaches to kiwifruit
Setting aside anti-herbivore or anti-pathogen roles that the fuzz-forming trichomes may play, a fuzzy specialized epidermis and cuticle appears to be one general strategy to avoid water loss or excessive heat and sunlight in xeromorphic plants, species with adaptations for tolerating sunny, dry environments (21). Sunny, semiarid conditions characterize many areas of especially western China, where both kiwifruit (2–4). and peaches were first domesticated (32).
Most of what we know about the function of fuzzy fruit surfaces comes from work on peaches, which are becoming something of a model organism for fuzz (25). The main function of those trichomes, as far as we understand it, is to help the peach avoid excessive water loss or sunburn (25). The hairs physically deflect hot sunlight and dry wind that would pull water away from the ripening peach and expose it to dangerous ultraviolet radiation. Trichomes therefore augment the water retention role of the wax-covered cuticle (25). The fuzz might also might make it more difficult for pathogens and pests to damage the fruit or seed, although this aspect his been less explored for peach (21, 25).

There is some evidence that kiwifruit trichomes also reduce desiccation. Golden kiwifruit was initially successfully commercialized alongside green kiwifruit in the mild, maritime climate of New Zealand. When golden kiwifruit was introduced into other regions of the world where green kiwifruits are commercially grown in hotter and drier conditions, such as California and Italy, the golden kiwifruits developed a shrivel disorder late in ripening, which is a failure of the plant vasculature to respond to sunny, arid conditions (33). Even when the irrigated vines themselves appeared healthy, the ripening golden kiwifruit lost water. Careful studies of water balance demonstrated that the cause was transpiration from the ripening fruit in excess of water inputs to the fruit from the vascular system (33). The hot, dry California environment simply forced the golden kiwifruit to lose more water than it did in cooler, more humid New Zealand, even though green kiwifruit ripened without issue in both environments (33). Specifically, the golden kiwifruit has a high surface conductance and transpiration rate compared with the green kiwifruit (33). Researchers confirmed that excessive water loss from the skin was the cause of the shrivel disorder in golden kiwi by conducting an experiment in which a plastic bag that increased the humidity of the air around the ripening fruit prevented the disorder (34).
I can think of two reasons why golden kiwifruit ripens less well in the golden state, which I’ll elaborate on below: (i) The trichome density of green and golden kiwifruits appears to be an adaptation to their respective environments of origin in different parts of China; and (ii) Commercial varieties of kiwifruits are especially close to their wild relatives, so we might expect high environmental fidelity for particular accessions.

Some fruit diversity in Actinidia, showing both fuzzy and smooth skin types (Fig. 1 from Crowhurst et al. 2008): “A is A. arguta, C is A. chinensis, D is A. deliciosa, E is A. eriantha, I is A. indochinensis, P is A. polygama and S is A. setosa.”
Kiwifruit trichome density reflects domestication history and polyploidy
As a formal agricultural enterprise, kiwifruit domestication is only about 100 years old (3) and is still very much underway. The most popular commercial varieties are only slightly removed from natural populations of Actinidia (2, 3, 35, 36), and new varieties are being identified from the wild and improved continually (recently, for example, A. eriantha; 37). Thus, commercial kiwifruit is quite similar to its wild relatives (3).
Golden kiwifruit (A. chinensis var. chinensis) naturally occurs mostly in the southern and eastern parts of China, in the subtropical, mild and wet locations in the Hunan Foothills. Green kiwifruit (A. chinensis var. deliciosa) occurs mainly in the western provinces, on the higher, more arid, Yun-Gui Plateau (2).
Polyploidy (multiple sets of chromosomes, as opposed to diploid, one copy) is rampant in in A. chinensis across its range (2–4, 36). Polyploid populations arise when duplicated genetic material is retained in a single gamete instead of being split into multiple gametes, and those gametes with extra genetic material nonetheless go on to produce viable offspring. Polyploidy is common in plants and is an important source of genetic variation for crop development (35). The A. chinensis varieties with the highest ploidy levels are found in the highest and driest spots (2). Populations of both green and golden kiwifruit from arid and high-altitude regions of China tend to be both polyploid and hairier.
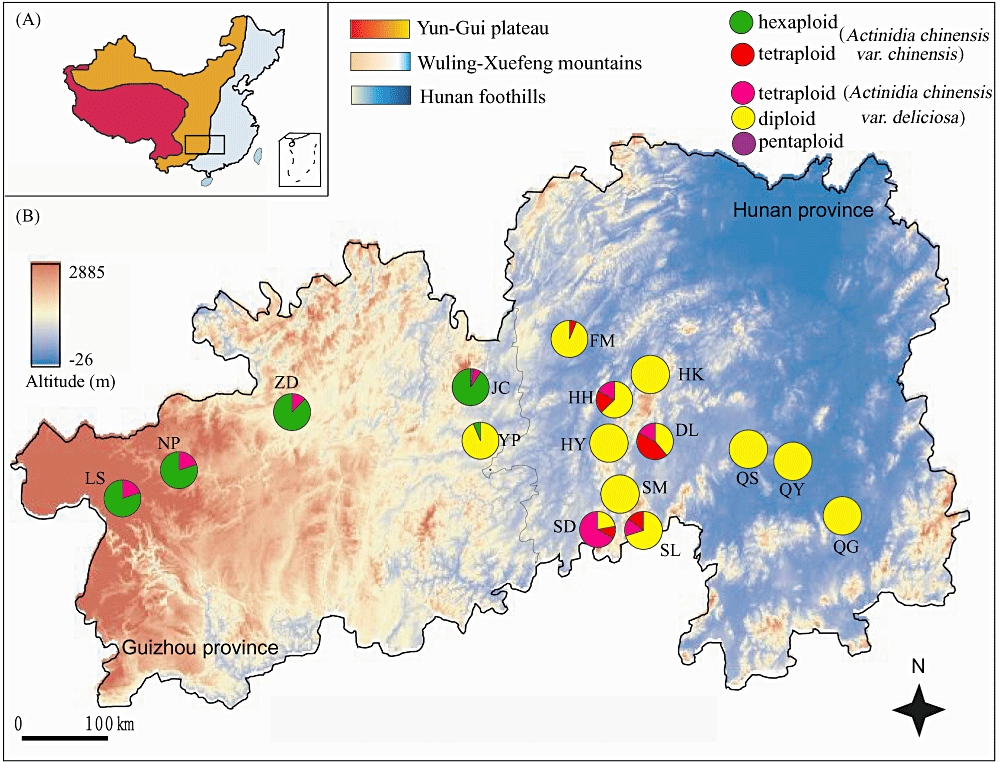
Progression of dominance from highly polyploid A. chinensis in arid western China to diploidy in the more moderate east, from (2): “The geographical distribution of 16 sample populations of Actinidia. A, The three steps of the Chinese topography are shown and the rectangle indicates the region in which the samples were obtained. B, Pie diagrams represent the proportion of ploidy races within each population. The colour spectrum of the map from blue to red indicates the altitude change.” I think the subspecies designation in the upper right corner here is misleading: all the hexaploid individuals in this study were var. deliciosa (very fuzzy green kiwifruit), and all the diploids were var. chinensis (less fuzzy golden kiwifruit).
Both polyploidy and increased trichome density (on at leaves if not fruit) have been positively correlated with the increased aridity and exposure to ultraviolet radiation typical of western China and high-altitude habitats in Actinidia (2, 3, 36, 38) and in some other taxa (39, 40). The Andean root vegetable maca is another example of a high altitude polyploid food plant. The most common variety of hairy green kiwifruit grown around the world, “Hayward,” is hexaploid (36), a polyploid with three copies of the genome (the ancestral state of Actinidia is diploid). All commercial hexaploid kiwfruit cultivars are green-fleshed, very hairy, and come from western China (36). Most commercial golden kiwifruit varieties are diploid, and therefore prone to less hairness and an eastern Chinese origin (36). The center of the ranges of most of the smooth-skinned kiwifruits overlaps with the highest incidence of diploid A. chinensis and slightly fuzzy golden kiwi (A. chinensis var. chinensis) in the relatively mild south- to central-eastern China (3), providing anecdotal evidence, at least, that trichomes aren’t necessary to prevent water loss in that more forgiving climate.

unpeeled, sliced kiwiberry (A. arguta)
More on peach vs. kiwifruit fuzziness
Katherine’s excellent essay on peach and nectarine genetics describes how changes in one gene ceases peach fuzz formation and turns the peach into a nectarine. While the mutant nectarines differ in many important respects from their peach progenitors, as far as I can tell, traits related to water loss or ultraviolet light avoidance have not yet been measured (41). Unlike with peaches and nectarines, we don’t know much about the genetic basis of trichome density in kiwifruit. We know that some genes are only expressed in fuzzy kiwifruits (42), and we know that all the non-fuzzy (glabrous) kiwifruit species are all in a single evolutionary clade (descendants from a single common ancestor; see our plant phylogeny page for a primer on these concepts), indicating that fuzzy fruit is the ancestral condition in Actinidia (43). Beyond that, though, there is obviously much work to be done to understand the genetic basis of fruit fuzziness.
Also, kiwifruit trichomes are multicellular, so they are a bit more complex than the single-celled trichomes of peach fuzz. If we infer that their function in preventing excessive water loss is similar, it begs the question of why peach produces simple, and thereby relatively cheap, trichomes, whereas kiwifruit produces more complex and therefore presumably more costly trichomes. Maybe simple trichomes wouldn’t do the job? Or the evolutionary capacity for one or the other types of trichomes simply wasn’t present? Much more work remains to be done on evolutionary pathways and selection pressures in trichomes.
The genomic (including ploidy) and genetic understanding of the characteristics that allow different fruit varieties to thrive in different environments is absolutely relevant to the production of fruit for human consumption. A plant can only breed with a compatible plant with the same ploidy level (see a good discussion of this in Katherine’s watermelon essay), so for kiwifruit, this affects the creation of hybrid lines in both the wild and on the farm, to produce desirable fruit that can thrive in different habitats. Reducing fruit hairiness is a target of kiwifruit breeding (3), but this effort will only be successful if the hairless fruits won’t shrivel in habitats where the vines supporting them otherwise thrive (as with the California-grown golden kiwi example).
Until that happy day of fuzz-less kiwifruit for all arrives, though, perhaps now it will be enough to have more food for thought while peeling.
References (below)
- Hallett IC, Sutherland PW (2005) Structure and development of kiwifruit skins. Int J Plant Sci 166(5):693–704.
- Li D, Liu Y, Zhong C, Huang H (2010) Morphological and cytotype variation of wild kiwifruit (Actinidia chinensis complex) along an altitudinal and longitudinal gradient in central-west China. Bot J Linn Soc 164(1):72–83.
- Huang H, Liu Y (2014) Natural hybridization, introgression breeding, and cultivar improvement in the genus actinidia. Tree Genet Genomes 10(5):1113–1122.
- Liu YF, Li DW, Huang H (2011) Cytotype variation in the natural populations of actinidia chinensis complex in China. Acta Horticulturae, pp 157–162.
- Dalin P, Ågren J, Björkman C, Huttunen P, Kärkkäinen K (2008) Leaf trichome formation and plant resistance to herbivory. Induced Plant Resistance to Herbivory (Springer Netherlands, Dordrecht), pp 89–105.
- Agrawal AA, Spiller DA (2004) Polymorphic buttonwood: Effects of disturbance on resistance to herbivores in green and silver morphs of a Bahamian shrub. Am J Bot 91(12):1990–1997.
- Hanley ME, Lamont BB, Fairbanks MM, Rafferty CM (2007) Plant structural traits and their role in anti-herbivore defence. Perspect Plant Ecol Evol Syst 8(4):157–178.
- Traw MB, Bergelson J (2003) Interactive effects of jasmonic acid, salicylic acid, and gibberellin on induction of trichomes in Arabidopsis. Plant Physiol 133(3):1367–1375.
- Atala C, Gianoli E (2009) Effect of water availability on tolerance of leaf damage in tall morning glory, Ipomoea purpurea. Acta Oecologica-International J Ecol 35(2):236–242.
- Sandquist DR, Ehleringer JR (2003) Population- and family-level variation of brittlebush (Encelia farinosa, Asteraceae) pubescence: Its relation to drought and implications for selection in variable environments. Am J Bot 90(9010):1481–1486.
- Arunprakash S, et al. (2014) Studies of Trichome in Five Species of Asteraceae. Int J Adv Interdiscip Res Sai Yaashitha Res Publ Int J Adv Interdis Res. Available at: http://ijaidr.co.in/files/Arun Prakash 1-4.pdf [Accessed November 2, 2017].
- Serna L, Martin C (2006) Trichomes: different regulatory networks lead to convergent structures. Trends Plant Sci 11(6):274–280.
- Agrawal AA, et al. (2009) Phylogenetic ecology of leaf surface traits in the milkweeds (Asclepias spp.): Chemistry, ecophysiology, and insect behavior. New Phytol 183(3):848–867.
- Sletvold N, Agren J (2012) Variation in tolerance to drought among Scandinavian populations of Arabidopsis lyrata. Evol Ecol 26(3):559–577.
- Shibuya T, Itagaki K, Ueyama S, Hirai N, Endo R (2016) Atmospheric humidity influences oviposition rate of Tetranychus urticae (Acari: Tetranychidae) through morphological responses of host Cucumis sativus leaves. J Econ Entomol 109(1):255–258.
- Kessler M, Siorak Y, Wunderlich M, Wegner C (2007) Patterns of morphological leaf traits among pteridophytes along humidity and temperature gradients in the Bolivian Andes. Funct Plant Biol 34(11):963–971.
- Karabourniotis G, Bornman JF (1999) Penetration of UV-A, UV-B and blue light through the leaf trichome layers of two xeromorphic plants, olive and oak, measured by optical fibre microprobes. Physiol Plant 105(4):655–661.
- Karabourniotis G, Fasseas C (1996) The dense indumentum with its polyphenol content may replace the protective role of the epidermis in some young xeromorphic leaves. Can J Bot 74(3):347–351.
- Ehleringer J (1982) The Influence of Water-Stress and Temperature on Leaf Pubescence Development in Encelia-Farinosa. Am J Bot 69(5):670–675.
- Ehleringer J, Mooney HA, Gulmon SL, Rundel PW (1981) Parallel Evolution of Leaf Pubescence in Encelia in Coastal Desert of North and South America. Oecologia 49(1):38–41.
- Skelton RP, Midgley JJ, Nyaga JM, Johnson SD, Cramer MD (2012) Is leaf pubescence of Cape Proteaceae a xeromorphic or radiation-protective trait? Aust J Bot 60(2):104–113.
- Yang Y, Korner C, Sun H (2008) The ecological significance of pubescence in Saussurea medusa, a high-elevation Himalayan “woolly plant.” Arct Antarct Alp Res 40(1):250–255.
- Vitarelli NC, Riina R, Cassino MF, Meira RMSA (2016) Trichome-like emergences in Croton of Brazilian highland rock outcrops: Evidences for atmospheric water uptake. Perspect Plant Ecol Evol Syst 22:23–35.
- Yang Q, et al. (2010) Research progress on water uptake through foliage. Acta Ecol Sin 30(3):172–177.
- Fernandez V, et al. (2011) New Insights into the Properties of Pubescent Surfaces: Peach Fruit as a Model. Plant Physiol 156(4):2098–2108.
- Fernández V, Khayet M (2015) Evaluation of the surface free energy of plant surfaces: toward standardizing the procedure. Front Plant Sci 6:510.
- Schreuder MDJ, Brewer CA, Heine C (2001) Modelled influences of non-exchanging trichomes on leaf boundary layers and gas exchange. J Theor Biol 210(1):23–32.
- Brewer CA, Smith WK, Vogelmann TC (1991) Functional interaction between leaf trichomes, leaf wettability and the optical properties of water droplets. Plant Cell Environ 14(9):955–962.
- Brewer CA, Smith WK (1997) Patterns of leaf surface wetness for montane and subalpine plants. Plant Cell Environ 20(1):1–11.
- Lusa MG, Cardoso EC, Machado SR, Appezzato-da-Glória B (2015) Trichomes related to an unusual method of water retention and protection of the stem apex in an arid zone perennial species. AoB Plants 7(1):plu088-plu088.
- Tsukaya H, Tsuge T (2001) Morphological adaptation of inflorescences in plants that develop at low temperatures in early spring: The convergent evolution of “downy plants.” Plant Biol 3(5):536–543.
- Li H-L (1968) The Origin of Cultivated Plants in Southeast Asia 1. Econ Bot 24(35):3–19.
- Clearwater MJ, Luo Z, Ong SEC, Blattmann P, Thorp TG (2012) Vascular functioning and the water balance of ripening kiwifruit (Actinidia chinensis) berries. J Exp Bot 63(5):1835–1847.
- Thorp TG, et al. (2007) “Hort16A” fruit beak end softening and shrivelling in California. Acta Horticulturae, pp 389–396.
- Li D, et al. (2014) Genetic diversity in kiwifruit polyploid complexes: Insights into cultivar evaluation, conservation, and utilization. Tree Genet Genomes 10(5):1451–1463.
- Li D, Zhong C, Liu Y, Huang H (2010) Correlation between ploidy level and fruit characters of the main kiwifruit cultivars in China: Implication for selection and improvement. New Zeal J Crop Hortic Sci 38(2):137–145.
- Wu YJ, et al. (2009) Characteristics of “white”: A new easy-peel cultivar of actinidia eriantha. New Zeal J Crop Hortic Sci 37(4):369–373.
- Liu Y, Liu Y, Huang H (2010) Genetic variation and natural hybridization among sympatric Actinidia species and the implications for introgression breeding of kiwifruit. Tree Genet Genomes 6(5):801–813.
- Guo W, et al. (2016) Divergence in Eco-Physiological Responses to Drought Mirrors the Distinct Distribution of Chamerion angustifolium Cytotypes in the Himalaya–Hengduan Mountains Region. Front Plant Sci 7:1329.
- Karl R, Koch MA (2013) A world-wide perspective on crucifer speciation and evolution: Phylogenetics, biogeography and trait evolution in tribe Arabideae. Ann Bot 112(6):983–1001.
- Wen I, Koch KE, Sherman WB (1995) Comparing Fruit and Tree Characteristics of Two Peaches and Their Nectarine Mutants. J Am Soc Hortic Sci 120(1):101–106.
- Kim SC, et al. (2004) Characterization of RAPD fragment unique to species with hairy fruit skin in the genus Actinidia. J Plant Biol 47(3):210–215.
- Huang H, Li Z, Li J, Kubisiak TL, Layne DR (2002) Phylogenetic Relationships in Actinidia as Revealed by RAPD Analysis. J AMER SOC HORT SCI 127(5):759–766.
- Guo X-M, Xiao X, Wang G-X, Gao R-F (2013) Vascular anatomy of kiwi fruit and its implications for the origin of carpels. Front Plant Sci 4:391.

Pingback: Bit fuzzy on kiwi fruit? – Agricultural Biodiversity Weblog
Pingback: Kiwifruit 2: Why are they green? | The Botanist in the Kitchen
Pingback: Evolution of Lemon Flavor | The Botanist in the Kitchen
Pingback: A Recipe for Botany | Mountain Plover
what an amazing post! it is both enjoyable and useful for my current project. I do thank you very much for all your research related with this topic 🙂
LikeLike
You’re welcome. Thanks for letting us know that the blog is working for you.
LikeLike
Pingback: Sage, rosemary, and chia: three gifts from the wisest genus (Salvia) | The Botanist in the Kitchen
Pingback: Kiwi Crush Paper: How a Fuzzy Fruit Became a Notepad - PaperPapers
Love this article! I was eating some home-grown kiwis and was curious about what the evolutionary point of the fuzziness is, and I couldn’t have asked for a better explanation; my only argument is that the skin is very much edible (and certainly if I tried to peel the ones I grew there’d be hardly anything left over 😆)
LikeLike