This essay is our annual contribution to the Advent Botany essay collection curated by Alastair Cullham at the University of Reading. We highlight three charismatic species in the large genus Salvia (in the mint family, Lamiaceae): rosemary, sage, and chia.
Two Christmases ago we pointed out the current fad in decorating pineapples for Christmas. This year, some of our gentle readers may come across potted rosemary bushes that are trimmed into a cone to resemble a conifer. These are pleasant and ostensibly can be kept alive after the holiday season.

A rosemary shrub trimmed into a conifer shape. Photo from Pottery Barn.
A perhaps less pleasant holiday botanical encounter may include a Christmas tree-shaped Chia Pet.

Christmas tree Chia Pet. Photo from Amazon.
As far as Chia Pets go, this one is fairly innocuous. In my view, however, its only saving grace is that the chia plant itself is a fabulous taxon (Salvia hispanica), as is the rest of its large genus, Salvia, which also happens to include rosemary (Salvia rosmarinus). Rosemary of course is much more likely to make a holiday appearance as a culinary ingredient than a decoration, lovely as it is. In the kitchen it is frequently joined with its congener Salvia officinalis, usually just called garden sage. That the genus Salvia is responsible for half the taxa in the title of a Simon & Garfunkel album (Parsley, Sage, Rosemary and Thyme), notwithstanding that Art Garfunkel looks like a Chia Pet on the cover, could provide enough taxonomic joy to justify leaving this examination of these plants here. The name “sage”, however, implies wisdom, and so like the wise men of old, I shall persevere.

Parsley, Sage, Rosemary and Thyme album cover by Simon & Garfunkel (1966)
We’ll start by addressing the taxonomic elephant in the room that might otherwise distract learned readers: rosemary was only brought into the Salvia fold in 2017. Before then it was in its own small genus: Rosmarinus. The reason Rosmarinus is now Salvia is that the speciose Salvia was found to be paraphyletic: the pre-2017 conscription of the nearly 1000 species in the genus did not include all of the descendants of their most recent common ancestor. When the relationships between all the Salvia species and their closest relatives were plotted on a single phylogenetic tree, it was obvious that Rosmarinus and a few other genera should more naturally be considered Salvia, and Salvia was revised accordingly.

Rosemary (Salvia rosmarinus)
Another taxonomic bookkeeping item is to clarify that the sages in Salvia are only distant relatives of the sagebrushes and sageworts in the genus Artemisia, which is in the sunflower family Asteraceae (please see our Artemisia essay for more information about that genus, which includes the herb tarragon). The phylogenetic relationships of the major groups in Salvia from the most recent revision (Drew et al., 2017) is shown below.

Figure 2 from Drew et al. (2017): “(A) Composite chronogram of subtribe Salviinae (which contains Salvia and related taxa) based on chloroplast DNA sequences from previous molecular phylogenetic analyses. Asterisks denote nodes with low support and/or conflicting resolution among previous analyses. Salvia nomenclature follows subgeneric clades described here, including three tentatively named clades that await proper circumscription. Calibrations based on Drew & Sytsma (2012; See supplementary figure S4) (B) Circle cladogram framed on larger chronogram with weakly supported nodes collapsed, depicting species diversity and generalized staminal types within each clade of Salvia; modified after Walker & Sytsma (2007) and Walker et al. (2015).” S. elegans (pineapple sage), S. sclarea (clary sage), and S. hispanica (chia) are in the American subgenus Calosphace. Rosemary is in its own subgenus, Rosmarinus.
The phylogenetic diagram above (from Drew et al., 2017) shows locations where the flower anther structure evolved into a lever-like mechanism that aids in bee pollination by physically moving the two stamens into contact with the bee’s back when a bee enters the flower (see illustration below from Walker, Sytsma, Treutlein, & Wink, 2004).

Figure 2 from Walker et al 2004: “Flower and pollination of Salvia pratensis (Salvia clade I). A flower without the lever mechanism activated (A). As the pollinator enters the flower (B), the pollen is deposited on the back of the pollinator. As the pollinator enters an older flower (stamens removed from sketch, but remain present in flower) pollen is transferred (C). The posterior anther thecae forming the lever can be fused or free and in the subg. Leonia, produce fertile pollen”
The lever mechanism independently evolved three times within Salvia. Each of these evolutionary events was followed by rapid and prolific speciation driven by this innovation in pollination biology (Drew et al., 2017): the advent of the lever mechanism led to the radiation of around 500 species in the subgenus Calosphace in Central and South America; around 250 species evolved soon after the advent of the lever mechanism in the Salvia officinalis clade in the Mediterranean and Western Asia; and around 100 species radiated following the lever in Far East Asia in the Salvia glutinosa clade.

Sage (Salvia officinalis) flowering on my deck this summer
The bee-pollinated Salvia flowers are distinct from those pollinated by hummingbirds, which are more elongate and often red, like the flowers of pineapple sage (S. elegans), and have either evolutionarily lost the staminal lever mechanism or never had it in the first place.

Pineapple sage (Salvia elegans)
The parsley, sage, rosemary, and thyme made famous by Simon & Garfunkel started their culinary careers in Europe. All but parsley are in the mint family (Lamiaceae; see our carrot top essay for a discussion of fun chemical relationships between the flavor compounds in the mint family and the parsley family, Apiaceae). This points to the profusion of aromatic mint family species common to the rocky shrublands covering much of Europe and western Asia (Rundel et al., 2016; Vargas, Fernández-Mazuecos, & Heleno, 2018).

Called “tomillar” in spanish, literally a field of wild thyme (Thymus vulgaris) and associated species growing in the Orusco de Tajuña hills (near Madrid. Spain). Other edible Lamiaceae can be found in this plant community, including Salvia rosmarinus, and Lavandula latifolia (a lavendar). Photo by Julia Chacón-Labella.
That broad area is one of the centers of Salvia species diversity, but the genus is globally widespread. The genus probably originated and dispersed first from African and then the Mediterranean (see the figure of Salvia distribution and putative dispersal history below from Will & Claßen-Bockhoff, 2017), but the full story of dispersal and species radiation within the genus requires more elucidation. Numerous species of Salvia are utilized as culinary or medicinal herbs or garden ornamentals throughout its range.

Fig. 8 from Will et al. 2017: “Salvia s.l. in time and space. A: Distribution of Salvia s.l., putative migration routes and fossil sites; BLB = Bering Land Bridge; D = Dorystaechas; M= Meriandra; NALB = North Atlantic Land Bridge; P = Perovskia; R = Rosmarinus; Z = Zhumeria; white arrows indicate repeated colonization of S Africa and dispersal from the Eastern Cape to Madagascar; hatched arrows (dark grey) indicate the repeated colonization of the Canary Islands from two different mainland sources; red arrow illustrate the dispersal from East Asia to Eurasia reflected by S. glutinosa; black arrows correspond to dispersal events from the OW to America reflected by two distinct lineages; ? = route uncertain; template of the map provided by the German earth science portal (www.mygeo.info). B: Simplified phylogenetic tree; nodes discussed in the text are indicated by capital letters; colors reflect distribution areas. (For interpretation of the references to color in this figure legend, the reader is referred to the web version of this article.)”
The phylogeny above shows the large number of American taxa in subgenera Calosphace and Audibertia. While many of these species have also been used as aromatic herbs and traditional medicines, the most famous of the American Salvias, chia, is known for its nutritious seeds (Jenks & Kim, 2013a). Chia is a name given to two species of Salvia: S. columbariae and S. hispanica. S. columbariae ranges from southern California to central Mexico, at which point the range of S. hispanica begins and extends to Guatemala. Indigenous groups throughout that range historically used both species of chia as a pre-Columbian staple food source. The Aztecs cultivated it, and 16th century Spanish codices indicate it may have been as widely utilized as maize (Cahill, 2003).

Chia nutlets (S. hispanica) and a dried sage (S. officinalis) leaf for scale
Technically, the chia “seeds” you can buy in the store (or harvest yourself) are fruits. The Salvia fruit, like those of all mint family species, is called a schizocarp. The ovary inside the flower has four chambers, called locules. Each locule matures into an independent, indehiscent nutlet. The shell (pericarp) of the nutlet is stratified into the same categories of outer fruit layers as are more familiar fleshy fruits (cuticle, epicarp, mesocarp, endocarp; see our pomegranate or apple essay for more details about fruit structure), but in the Salvia nutlet the outer fruit layers are dry and compressed and inseparable from the single seed inside the fruit (Capitani, Ixtaina, Nolasco, & Tomás, 2013). Salvia nutlets mature inside of papery fused calyces (see the photo below of sage nutlets and their cup-like persistent calyces).

Sage (Salvia officinalis) leaves and nutlets inside of papery, fused persistent calyces.
The word “chia” is derived from the Aztec language Nahuatl word for “oily,” a name bestowed because chia seeds do have a high oil content (Cahill, 2003). Chia oil is rich in the omega-3 fatty acid alpha-linolenic acid, which has contributed to its recent fame as a modern health food. High alpha-linolenic acid content may be a general feature of the genus: other Salvia species, including S. officinalis, garden sage, have been shown to have high alpha-linolenic acid content in their seeds (Ben Farhat, Chaouch -Hamada, & Landoulsi, 2015).
Chia nutlets are also known for the gooey mucilage they exude when wet. This polysaccharide matrix is used as a food binder and thickener (Google “vegan egg replacement”). The production of mucilaginous diaspores (the dispersing agent, a fruit or a seed) is called myxocarpy. As Katherine discusses in her essay on okra, the flagship mucilaginous food plant, the purpose of the mucilage is likely water retention in the arid regions where these plants tend to come from. The mucilage might also act as a glue to bind the nutlet to the soil or to a dispersing animal’s fur—or to the terracotta substrate of a Chia Pet. Myxocarpy is most common in plants with small seeds growing in dry, arid areas, like those where Salvia species have radiated (Ryding, 1992).

Sage growing in coastal California, a Mediterranean-type ecosystem
Within the mint family, myxocarpy only occurs in the subfamily Nepetoideae. The subfamily, incidentally, gets its name from the catnip genus, Nepeta. Most if not all of the familiar edible herbs from the mint family are in this subfamily. Katherine has taken advantage of myxocarpy in this clade by serving soaked black basil (Ocimum basilicum) nutlets as a basil-scented vegan “caviar.”

Cat in the catnip (Nepeta cataria)
Salvia aroma and flavor–and I think the psychoactive properties of catnip for cats and known hallucinogen Salvia divinorum–comes from the terpenoids and phenolics that comprise their essential oils. The terpenoids are synthesized and stored in special glandular trichomes on the leaf surface (Schuurink & Tissier, 2019). Trichomes are hair-like extensions of the epidermis, although the glandular trichomes full of essential oil look more like water balloons than hair. Salvia species have other types of trichomes in addition to the glandular trichomes that are indeed much more hair-like and give the leaves of some Salvia a downy or prickly appearance (Kamatou et al., 2006).
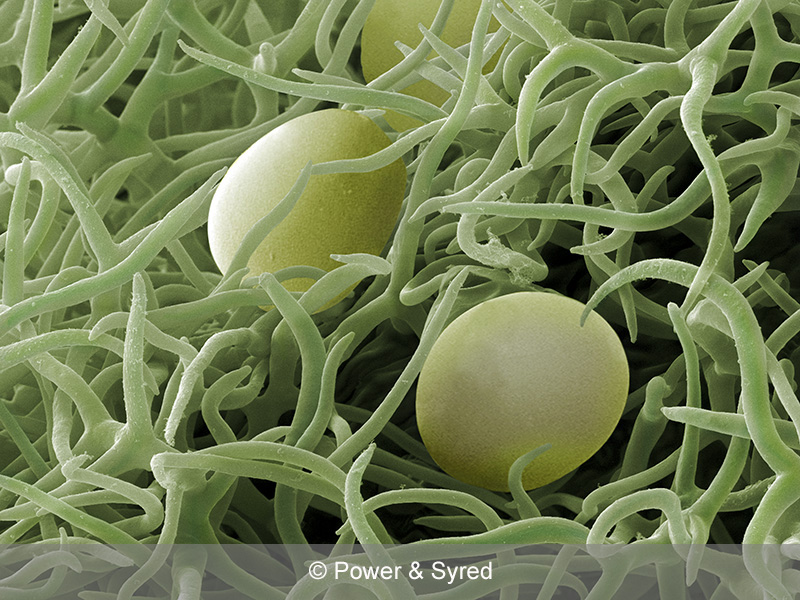
Scanning electron micrograph (SEM) of a rosemary leaf. Spherical oil-filled glandular trichomes are found amongst the branched hair-like trichomes covering the lower surface of the leaf, which has a greater profusion of hairs and glands than the upper surface. When the glands are damaged or broken the aromatic essential oil is released. Magnification: x1550 (x381 at 10cm wide). Photo from https://psmicrographs.com/sems/flowers-plants/
We discussed trichome function extensively in one of our kiwi essays. The hair-like trichomes may serve the leaf by protecting it from excess solar radiation and wind and otherwise creating a more mild microclimate at the leaf surface to help it retain water.

Rosemary
Terpenoid biosynthesis requires numerous steps in which intermediate chemical products are modified by a series of specific enzymes and other proteins. Small changes in the genes responsible for those proteins can lead to big qualitative changes in the final terpenoid mix in the essential oil of a given taxon. We mammals are adept at discerning aroma differences between chemically similar terpenoids. For example, in on our carrot top essay we discussed the case of spearmint and caraway. The respective versions of the terpenoid carvone that characterize the essential oils of those plants differ only in the physical configuration of the same chemical elements, but they smell radically differently to us.

Clary sage (S. sclarea)
The function of the essential oil in the glandular trichomes, however, is not to improve human well being. Plants synthesize those lovely terpenoids as chemical defense against insect herbivores and microbial pathogens. When the hair-like trichomes fail to stop the intruders, the glandular trichomes will explode on contact, drenching the would-be attackers in a caustic-but-fragrant deluge.

rosemary
The pharmacopeia of terpenoid aromas present in the mint family—bring to mind the scents of sages, rosemary, lavender, peppermint, spearmint, savory, thyme, oregano, marjoram, shiso, basil—owes its evolutionary origins certainly in part at least to the various selection pressures imposed on those herbal taxa by their pests. Within even commonly grown domesticated Salvia species, essential oil constituent variation leads to dramatic differences in aroma. For example, consider the differences among rosemary, garden sage, clary sage (S. sclarea), and pineapple sage (Salvia elegans), which has a notably fruity smell. The fruitiness is due in part to the presence of the terpenoids charcteristic of citrus, which are widespread across plants.

Garden sage (Salvia officinalis)
The Roman historian and natural scientist Pliny the Elder coined the name Salvia, which is derived from the Latin salvare, meaning to heal and save, and salvus, meaning uninjured or whole. The common English name “sage” of these plants ultimately comes from this same Latin root. In Pliny the Elder’s time, the Mediterranean Salvia species were considered healing herbs, good for treating colds and a variety of ailments. Salvia feature prominently in the ethnomedicine of every region in which it is found (South Africa: (Kamatou et al., 2006); Central and South America: (Jenks & Kim, 2013b)). There is a Chinese proverb that asks “How can a man grow old who has sage in his garden?” I do not know which Salvia species would have been responsible for this proverb. There are over a hundred species of Salvia species native to China, and the Mediterranean import Salvia officinalis is grown throughout the country.

Bundle of dried sage, recently, recently, in Alaska
The health and wellness meaning of “sage” is etymologically independent from its other definition as a wise thing or wise person. This second meaning ultimately comes from the Latin sapere, to know or taste. I personally enjoy conflating these meanings, tying wisdom and well-being to the plant. I like that the Salvia officinalis that grew on a pot on my deck this summer and that will season comfort food this winter is a descendent from the plants that healer contemporaries of Pliny the Elder would have searched for amidst sun-drenched rocks in the Mediterranean hills.

Salvia in macarons at my local bakery (Fire Island) this week: blackberry-sage and rosemary-merlot.
Simon & Garfunkel close the Parsley, Sage, Rosemary and Thyme album with the song “7 O’Clock News/Silent Night,” in which they juxtapose jarring newscasts from the Nixon and Johnson era with the Christmas carol. This holiday season has felt a bit like that song to me, like concerted effort is required to prevent awful, omnipresent news from drowning out the joy and solemnity of marking the darkest time of the year. But perhaps honoring traditions always involves this element of deliberately carving out the space in which to do so. Perhaps sprinkling rosemary and sage into a holiday stew or stuffing can be a radical act, a defiant embrace of old wisdom to fortify ourselves to stand with each other and create something beautiful in the cold. Regardless, insane amounts of butter will be involved, at least at my house. And when the January 2nd resolutions to “eat better” come around, chia will be there.
References
Ben Farhat, M., Chaouch -Hamada, R., & Landoulsi, A. (2015). Oil yield and fatty acid profile of seeds of three Salvia species. A comparative study. Herba Polonica, 61(2), 14–29. doi:10.1515/hepo-2015-0012
Cahill, J. P. (2003). Ethnobotany of Chia, Salvia hispanica L. (Lamiaceae). Economic Botany, 57(4), 604–618. doi:10.1663/0013-0001(2003)057[0604:EOCSHL]2.0.CO;2
Capitani, M. I., Ixtaina, V. Y., Nolasco, S. M., & Tomás, M. C. (2013). Microstructure, chemical composition and mucilage exudation of chia ( Salvia hispanica L.) nutlets from Argentina. Journal of the Science of Food and Agriculture, 93(15), 3856–3862. doi:10.1002/jsfa.6327
Drew, B. T., González-Gallegos, J. G., Xiang, C. L., Kriebel, R., Drummond, C. P., Walker, J. B., & Sytsma, K. J. (2017). Salvia united: The greatest good for the greatest number. Taxon, 66(1), 133–145. doi:10.12705/661.7
Jenks, A. A., & Kim, S. C. (2013a). Medicinal plant complexes of Salvia subgenus Calosphace: An ethnobotanical study of new world sages. Journal of Ethnopharmacology, 146(1), 214–224. doi:10.1016/j.jep.2012.12.035
Jenks, A. A., & Kim, S. C. (2013b). Medicinal plant complexes of Salvia subgenus Calosphace: An ethnobotanical study of new world sages. Journal of Ethnopharmacology, 146(1), 214–224. doi:10.1016/j.jep.2012.12.035
Kamatou, G. P., van Zyl, R. L., van Vuuren, S. F., Viljoen, A., Figueiredo, A. C., Barroso, J. G., … Tilney, P. M. (2006). Chemical composition, leaf trichome types and biological activities of the essential oils of four related Salvia Species indigenous to Southern Africa Analysis of plant volatile using 2D gas chromatography View project Chemometrics View project. Journal of Essential Oil Research. Retrieved from https://www.researchgate.net/publication/236850867
Rundel, P. W., Arroyo, M. T. K., Cowling, R. M., Keeley, J. E., Lamont, B. B., & Vargas, P. (2016). Mediterranean Biomes: Evolution of Their Vegetation, Floras, and Climate. Annual Review of Ecology, Evolution, and Systematics, 47, 383–407. doi:10.1146/annurev-ecolsys-121415-032330
Ryding, O. (1992). Pericarp structure and phylogeny within Lamiaceae subfamily Nepetoideae tribe Ocimeae. Nordic Journal of Botany, 12(3), 273–298. doi:10.1111/j.1756-1051.1992.tb01304.x
Schuurink, R., & Tissier, A. (2019). Glandular trichomes: micro-organs with model status? The New Phytologist, nph.16283. doi:10.1111/nph.16283
Vargas, P., Fernández-Mazuecos, M., & Heleno, R. (2018). Phylogenetic evidence for a Miocene origin of Mediterranean lineages: species diversity, reproductive traits and geographical isolation. Plant Biology, 20, 157–165. doi:10.1111/plb.12626
Walker, J. B., Sytsma, K. J., Treutlein, J., & Wink, M. (2004). Salvia (Lamiaceae) is not monophyletic: implications for the systematics, radiation, and ecological specializations of Salvia and tribe Mentheae. American Journal of Botany, 91(7), 1115–1125. doi:10.3732/ajb.91.7.1115
Will, M., & Claßen-Bockhoff, R. (2017). Time to split Salvia s.l. (Lamiaceae) – New insights from Old World Salvia phylogeny. Molecular Phylogenetics and Evolution, 109, 33–58. doi:10.1016/j.ympev.2016.12.041
