With so many strawberries in so many kitchens this month, now is the perfect time to merge botany lab and breakfast preparation by working through the many parts of a strawberry. We kick off the strawberry love with some explanations in Food for Thought. You can learn quite a bit about flowers by cutting up a strawberry, as we do in this month’s Botany Lab of the Month. As it turns out, this enigmatic little gem is packed with coincidences and apocrypha along with its citric acid and anthocyanins. We conclude with recent relevant media and journal articles in Gleanings and Baker’s Dozen.
Developing coconuts, in Panama. Photo by J. Touchton
The May 2026 issue of our Gather and Sow newsletter is out.
Coconut palms grow some of the biggest seeds on the planet, and vanilla seeds, from an orchid, are among the smallest. They are both delicious! They are also the subjects of this month’s newsletter. Just in time for Mother’s Day (May 10th this year), in this month’s Food for Thought essay (“Coconuts for Mother’s Day”), we make the case that Mother’s Day celebrations should include more coconut. The large seed size of coconuts, and the small size of orchid seeds, are indicative of their respective parenting philosophy. In this month’s Botany Lab of the Month we describe coconut and vanilla fruit and seed morphology in the course of making cookies (“Extreme Monocot Cookies”). The newsletter concludes with Gleanings and Baker’s Dozen, our roundup of relevant recent media and journal articles.
The Gather and Sow: April 2026 newsletter is out, just in time for Easter. We write about betalains, the pigments that impart brilliant colors to beets and their relatives. And we dye Easter eggs with plant dyes, make violet syrup, and learn about pH.
“Light Iris” by Georgia O’Keefe (1924). You don’t have to seek out a Georgia O’Keefe painting to see giant flower parts from the iris family (Iridaceae). Just look in your spice cabinet. The spice saffron is the dried stigmas from the flowers of the saffron crocus (Crocus sativus), also in the iris family. We explore the botany of saffron and corn silks in this month’s Botany Lab of the Month feature.
Leeks are a traditional symbol of St. David’s Day, and their morphology is the topic of this issue’s Food for Thought: Ay, leeks is good!
Women’s history month is a time to recognize the contributions of women to American history, society, and culture. Too often women’s accomplishments have gone unsung. The same is true for the “female” parts of flowers. We rectify this by celebrating extraordinary flower parts in corn and the saffron crocus. All celebrations deserve cake, and we offer a recipe for one that combines corn and saffron in this month’s Botany Lab of the Month: Honoring female (plant) anatomy with saffron polenta cake.
We conclude with research and media highlights in Baker’s Dozen and Gleanings. Alongside several recent research articles, we have included a few classic papers by two matriarchs of plant biology, Katherine Esau and Barbara McClintock, in honor of Women’s History Month.
Cauliflorous fruits on a cacao tree (Theobroma cacao), in a glasshouse at the United States Botanic Garden.
We intend to post new issues of Gather and Sow every month. Content here on the blog will generally remain available. We remain dedicated to illuminating the fascinating biology of our food plants.
Each issue of Gather and Sow will feature three components:
Food for Thought: an essay, possibly seasonal or inspired by current events
Botany Lab of the Month: a botany lesson for the hands-on learner, packaged as an activity or recipe designed for the home kitchen
Baker’s Dozen and Gleanings: We highlight thirteen-ish recent research articles and other relevant media, respectively, that further our understanding of the biology of our food plants.
FAQ: Aren’t you two writing a book? Is it done yet? Yes, we are writing The Botanist in the Kitchen book. No, it is not done yet. We have an agent (Hi Lucy!). And a book proposal document. We are close to being done with our sample chapters. Hopefully a publisher will take interest very soon. We’ll post updates with the newsletters.
We would love to hear from you.
You can subscribe to Gather and Sow from theGhost site. We will also publicize newsletters here on the blog and on social media (Bluesky, Facebook). Please email us at botanistinthekitchen@gmail.com.
A Seminole pumpkin in my garden near DC, a decade ago
A Seminole pumpkin in a CSA box a decade ago got me thinking about squash diversity. Market bins and seasonal displays that autumn overflowed with squashes, gourds, and pumpkins in a dizzying array of sizes, shapes, and colors, but I had never seen a Seminole pumpkin among them. My pumpkin was squat, round, and the size of a cantaloupe. Its smooth, matte tan skin exactly matched the exterior of a butternut squash. The dense, dark orange flesh in its interior matched, too. It turns out that there is a good reason for this similarity: butternut squashes and Seminole pumpkins are different varieties of the same species, Cucurbita moschata. The pureed squash inside a can of commercially canned pumpkin is yet another variety of C. moschata, the Dickinson pumpkin, developed in the early 1800s by a Kentucky farmer named Elijah Dickinson. C. moschata boasts numerous other varieties, names of which variously include “squash” or “pumpkin”, but it is rarely the most well represented squash species at the market. That honor usually goes to Cucurbita pepo, followed closely by Cucurbita maxima, two of the five species of Cucurbita whose fruits appear on our tables as tender-skinned summer squashes or hard-shelled winter squashes.
Squash origins
All Cucurbita species are native to the Americas, a dozen or so species with scattered wild distributions, mostly in Mexico. Cucurbita fruits were important dietary staples for indigenous peoples from Central America to New England. Domestication of at least five Cucurbita species predated European exploration of the Americas by several thousand years. C. pepo may have been domesticated in Mexico by ten thousand years ago, around the same time that wheat was domesticated in the Mediterranean.
The word “squash” is derived from askutasquash, meaning “that which is eaten raw or uncooked” in the Algonquian language Narragansett, spoken by some indigenous groups of northeastern North America. It may seem surprising to think of winter squash as raw or uncooked, but drying strips of the raw fruit was a common means of traditional preparation and preservation throughout the Americas. The Massachusett and Wampanoag peoples of New England had an additional word, pôhpukun, to describe the squashes they grew that “grow forth round.” This word was transmogrified into “pumpkin” by the 17th-century English colonists made famous by tales of the first Thanksgiving. The indigenous word was conveniently similar to the Early Modern English word for Eurasian melon, pompion, which was familiar to Europeans of the period.
Sugar pumpkins (C. pepo) in a market
C. moschata proved particularly hardy in the swampy lands of what is now the southeastern United States, traditionally home to numerous tribes, including the Seminole. The squash that now bears their name grows as a sprawling plant with the huge palmate leaves and curling tendrils typical of cucurbits, members of the gourd family (Cucurbitaceae), which also includes cucumbers, melons, and gourds (and loofah!). Accounts of early Spanish explorers to Florida describe Cucurbita vines twining up tree trunks, with their pendulous fruits hanging over rivers or decorating the dead oaks that the Native Americans used as trellises to farm the squashes. The large nutritious seeds inside the fruit were as important a food source as the sweet flesh. Unsliced fruit could be stored for months, protected by the hard outer shell.
Seminole pumpkin (C. moschata) vining up my house
Squash hardiness is usurprising given the means by which cucurbit lineages initially arrived in the Americas. The family Cucurbitaceae arose near modern-day India in the late Cretaceous, around 63 million years ago. The leading hypothesis for how ancient cucurbits migrated from the Asian subcontinent to every other continent except Antarctica is transoceanic long-distance dispersal. That is, either the tough-shelled fruits floated across the ocean, or the seeds were carried in the guts of birds. Ancestral cucurbits made the journey from Asia to Africa, and then from Africa to South America. The jump from Africa to South America happened approximately five times over the course of several million years. The descendants of those five founders eventually radiated into around 350 modern cucurbit species in the Americas. The genus Cucurbita originated between 9 and 23 million years ago in Central or South America. Cucurbita expanded its range into North America by the onset of indigenous American agriculture, around 10,000 years ago.
Five species, many varieties
Butternuts (C. moschata) in a market
Those northern radiations of the genus resulted in the three highly variable species that constitute the vast majority of pumpkins and other squash consumed in the world: Cucurbita pepo, C. moschata, and C. maxima. Two additional Cucurbita species, C. argyrosperma (cushaw) and C. ficifolia (fig-leaf gourd), are cultivated in a few areas, mostly in Mexico and Central America. Most of the squashes that come to mind when you think of “pumpkin”—large Jack-O-Lantern pumpkins, orange sugar pumpkins, squat mini pumpkins, giant pumpkins, white Casper pumpkins—are varieties of Cucurbita pepo. C. pepo also includes acorn squash, delicata, stringy spaghetti squash, and most of the summer squashes—zucchini, yellow crookneck, pattypans, and marrows. C. maxima includes the hubbard, turban, and kabocha squashes. Distinct named cultivars of these Cucurbita squash species have been developed through selective breeding over centuries, and most are totally interfertile within a species, and the species do occasionally hybridize.
Summer squash cultivars–especially the zucchini, yellow crookneck, and vegetable marrow cultivars of C. pepo–have been developed for their tender, immature fruit. These fruits are meant to be picked well before seed maturation. If left on the plant to mature, summer squashes will develop the hard rind and woody seeds that characterize their winter squash brethren. European explorers introduced hard-shelled American Cucurbita to the rest of the world starting in the early 16th century, and the agricultural development of summer squashes occurred primarily in Europe subsequently. Most squash varieties, however, have been developed under selection for various characteristics of the mature fruit. At least one variety in Mexico was developed primarily for the mature seeds—pepitas—a staple of Oaxacan cuisine.
Much is known of the genetic basis of the tremendous morphological variation among Cucurbita species. We largely understand the genetic architecture of fruit shape, shell lignification (becoming hard and woody), color, size, and beta carotene content (carotenes—Vitamin A precursors—make the fruit of most winter squashes yellow to orange). A white pumpkin, for example, expresses dominant alleles (gene variants) of two genes: Wf for white flesh and W for weak fruit color. A warty pumpkin has a dominant allele of the gene Wt. The traditional orange pumpkin has the right alleles for many genes responsible for the synthesis of orange carotenoid pigments, especially lutein and beta-carotene, and the “orange” gene Or encodes an enzyme that directs the differentiation of specialized plastids in fruit cells called chromoplasts in which those carotenoids accumulate.
Summer squashes (C. pepo)
A particularly interesting gene unique to C. pepo called sp controls the stringiness of the fruit. When a hapless C. pepo inherits two copies of the recessive allele (gene variant) of the sp gene from its parents, the fruit flesh will have “spaghetti” texture, breaking into long strands when cooked. Hence, the name for “spaghetti squash” or “vegetable spaghetti.” Anatomically the strings are separated by bands of pectin that disintegrate during cooking. As cultivars of C. pepo, Jack-O-Lantern pumpkins, acorn squash, and over-ripe zucchini can also be stringy. If you cook up your Jack-O-Lantern pumpkins after Halloween, you might have to put the flesh through the blender to puree the strings. The flesh of C. maxima and C. moschata tends to be firmer than that of C. pepo, and it cooks up smoother. This is undoubtedly why C. moschata is preferred as the “pumpkin” of choice for commercial canned pumpkin producers, and why I prefer C. moschata or C. maxima varieties for any recipe that calls for pureed squash, be it ravioli or pumpkin pie.
Turning pumpkin carving into a botany lab
Amish pie pumpkin, a large and delicious variety of hubbard squash, Cucurbita maxima. That’s a 12-inch chef’s knife for scale.
Winter squashes are gloriously large fruits and are therefore excellent subjects for botanical observation. Carving a Jack-O-Lantern or cutting up a squash for a recipe becomes an instructive dissection when you know what to look for.
Botanists have names for different types of fruits. A squash is a pepo, a hard-rinded berry that develops from a flower with a single inferior ovary. “Inferior” means that the squash flower ovary is located within the hypanthium, the tissue that supports the petals and sepals. Squash plants make separate male and female flowers. Upon opening, the golden petals of female flowers are already subtended by a tiny squash—the inferior ovary and its protective tissue. The round brown circle on what we perceive to be the bottom of the pumpkin is the scar left after the petals and sepals fell off the developing fruit. As the fruit develops, the hypanthium tissue fuses with the outer wall of the ovary to become the exocarp rind, encasing the firm flesh of the mesocarp and soft endocarp. When you scoop out the stringy, gooey stuff and seeds from inside the pumpkin, you’re scraping out both the endocarp and the placental tissue that that connects the seeds to the ovary wall. The cavity of your pumpkin might look like it has lobes to it, called locules. I start the pumpkin carving by cutting out the lid around the “handle,” which is really the peduncle, the specialized stem that connects first the flower and then the developing fruit to the main plant stem.
Save the seeds! It’s a bit of a pain to separate the gooey fruit from the seeds, but salted and roasted pumpkin seeds is a real treat. Crack the hard seed coat (testa or integument) to reveal the olive-green-colored cotyledons (leaves) of the embryo, which take up most of the seed. The part of the embryo that will sprout roots is in the pointy end of the teardrop-shaped seed.
Seeds of ancestral Cucurbita species are present in fossilized mastodon dung deposits, which suggests that fruits of the genus were already adapted to dispersal by large mammals even before humans got involved with its evolution through domestication. There is evidence that some ancient Cucurbita species declined in geographic extent and abundance following the extinction of the animals that had pushed the evolution of its fruits toward large size and high sugar content. Humans arriving in the Americas essentially replaced the extinct megafauna and fundamentally changed the trajectory of the languishing Cucurbita, which in turn eventually became a crucial food source for the indigenous peoples of two continents.
I find this long view of the history of these plants and people to be a hopeful tale. It is as good an accompaniment to a Thanksgiving pumpkin pie as the whipped cream.
Where have the botanists in the kitchen been? In our writing rooms, working on other ways to bring you this content. We can’t wait to share what we’ve been doing!
Meanwhile, we hope you enjoy our substantial archive and follow us on Bluesky (@bitkblog.bsky.social, katherineapreston.bsky.social, jldosnas.bsky.social)
It’s the winter holiday season, when halls are bedecked with garlands of evergreens, sprigs of holly, and bunches of mistletoe to remind us that there is life in the darkness and love to be shared. This year, Katherine has added another symbolic plant to her own holiday list – pine nuts. They are more precious this year than ever.
I first started using pine nuts in holiday baking for the simple reason that they taste like pine and thus add a Christmas-tree note that almonds do not. A deeper significance was not on my mind. But pine nuts are so interesting botanically that I always slice some of them open to look for tiny pine embryos inside, and that triggers some nostalgia for conifer week in the botany lab I taught as a graduate student (along with the best co-TA ever). Specifically, I think about an odd conversation with one of the students, which for years was nothing but another funny story. Only now, decades later, do I understand that this student’s observations have something to teach us about the true meaning of pine nuts.
The remarkably unfiltered conversation happened after our student, while dissecting a pine nut, had experienced a double epiphany: he finally understood the details of sexual reproduction in pines, and he therein discovered a pathetically apt metaphor for his love life. I can still see the way he dropped his shoulders as dejection slid across his face. His exact words are lost after so many years, but he basically confided to us that, like a pine seed, he always invested a little too much and a little too early in the promise of love (or at least sex) which might never be fulfilled.
Lessons from pine sex
Both pines and flowering plants make seeds, however they don’t feed their embryos the same way. Pines (and other gymnosperms) pack a fat lunch in anticipation of an embryo, whereas flowering plants typically wait for successful fertilization and only then build up a food reserve for the embryo. Pines invest in an uncertain future, while flowering plants hold back and hedge their bets. Our student thought that his was a losing strategy, and that he should behave more like a flowering plant, but I’m not sure. I like to imagine that someone found his earnest vulnerability charming, and that he has found the loving partnership he was looking for. No matter what happened in his case, however, this I now know for certain: sometimes in life you have to muster the courage to invest fully, even recklessly, in hope. I think that’s a pretty good message for the short days of winter.
The full story of pine reproduction starts with the story of seeds, which is very complicated and still not fully resolved, but here it is in a nutshell. Seeds were an incredibly successful evolutionary innovation because they took a process that depended on wet soil or water pooling on bark or in sidewalk cracks and brought it inside a protective shell that remained on the parent plant and could function without free water. The ancestors of seed plants were similar to today’s ferns, in that they shed spores that germinated in moisture and grew into tiny plants that made eggs and swimming sperm. There are variations on this basic system throughout the plant kingdom, but ferns are a familiar example. In ferns, the large frondy generation is called the sporophyte (“spore plant”) because it makes spores. (Spores result from meiosis, so they contain half as many chromosomes per cell as the sporophyte does). Spores germinate and grow into flat green plants about the size of a lentil. These are called gametophytes because they make gametes (eggs and sperm). Under the right conditions, eggs and sperm meet, and the result is a new sporophyte.
The tree in the center is a flat-topped Italian Stone Pine (Pinuspinea) on the Stanford campus
Pine trees are also sporophytes, but they hold onto their female spores, which develop into egg-producing female gametophytes* inside the seeds on the scales of their cones. The male spores are shed as pollen grains, with sperm-producing cells inside. Whereas free-swimming fern sperm cells get nowhere without a film of water between themselves and some eggs, pine sperm packaged into pollen grains can float through the air towards more distant eggs. Although non-seed plants have done well evolutionarily – mosses and ferns are especially diverse, widespread, and abundant – the seed habit has freed gymnosperms and angiosperms from some ecological constraints and has undoubtedly contributed to their success in a range of habitats.
What is a pine nut?
For all its oily goodness, botanically a pine nut is not a nut at all. It is a seed, and without its shell (the seed coat), a pine nut is essentially nothing but female gametophyte, often with a cute little embryo inside bearing tiny pine needles. Long before it gets to that point, however, the gametophyte has to do what its name calls for – it makes eggs, two of them – and it also accumulates a lot of nutrients for a potential embryo. A typical commercial pine nut is about two-thirds fat by weight and one-third protein and carbohydrates. A tree invests in making hundreds or thousands of those energy-rich structures each season, even though only some of the eggs will be fertilized. I don’t know what proportion of the ovules (immature seeds) are actually fertilized on a typical tree, but in a bag of pine nuts it is sometimes hard to find any with an embryo inside. Other times, most of the seeds I open up do contain baby pines.
Pine nuts are worth dissecting in your kitchen because they give you a rare glimpse into the evolutionary history outlined above. By contrast, you will never see the female gametophyte of a walnut, pecan, almond, hazelnut, peanut, or cashew, at least not in your kitchen. In flowering plants, the female gametophyte has evolved to be just a handful of cells, and when we eat an angiosperm seed, we are eating some combination of embryo and that special made-just-in-time tissue called the endosperm.
Conifer week in your kitchen
Just for auld lang syne, I gathered and photographed some of the materials we might have used during conifer week in botany lab so that you can follow along at home. If you have your own pine nuts, that’s even better. Epiphanies are encouraged but not required.
Commercial pine nuts are harvested from natural stands of a few large-seeded species. European pine nuts come from the Italian stone pine, Pinus pinea, which is planted as an ornamental in other Mediterranean type climates, including, fortunately, the campus of Stanford University where I teach. Squirrels are also a conspicuous part of the flora and fauna at Stanford, and they had already taken most of the seeds out of the cones that I picked up. In fact, pine nut processors usually harvest cones directly from the tree before their scales have opened up, and the cones are allowed to dry at a safe distance from seed predators. Unfortunately, the remaining seeds I found, stuck inside their cones and spurned by the squirrels, had become moldy, so all the photos here of gametophytes and embryos are from pine nuts I bought. Those were harvested in China and came from a different species, the Korean pine (Pinus koraiensis).
Seed poking out from between cone scales
For most of their development – between pollination and seed release – pine cones keep their scales tightly closed. You can usually find cones in various stages of development on a tree because the whole process can take two or three years. When seeds are mature, the scales of most species open up, and the seeds can be seen peeking out from between them.
In pine species with small seeds, there is a prominent wing on each seed, and seeds flutter out away from the parent plant. Italian stone pines have very large seeds whose useless vestigial wings detach from the seeds easily.
The “shell” of a pine nut is nothing but a hard thick seed coat, its only protection against the outside world. This is what it means to be a gymnosperm — a naked seed. By contrast, the shells of other “nuts,” like pecans, almonds, or pistachios are part of the angiosperm fruit wall that surrounds the seeds, and their seed coats are very thin.
Most pine nuts are sold as bare gametophytes, without their seed coats. If you look at their pointed tips you can see a small opening where the pollen grains would have settled in to germinate and send out their pollen tubes. Pine gametophytes make two eggs in special chambers (archegonia), but usually the first egg to be fertilized is the only one that ultimately develops. I have never found twin embryos inside a pine nut, but it does happen. Twinning can also result when one embryo splits lengthwise early in development.
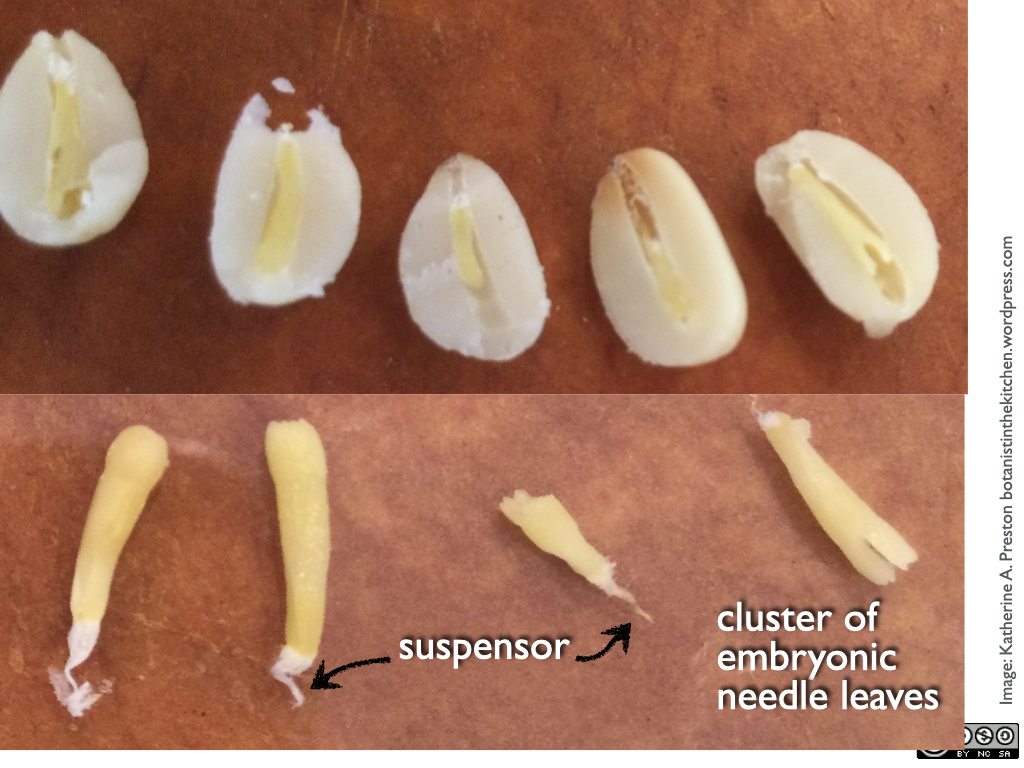
If you have any pine nuts to dissect, it’s best to use a razor blade because kitchen knife blades are a little too thick to do the job without mangling the embryo. A longitudinal section starting at the pointed tip reveals the embryo inside.
Above: row of pine gametophytes with embryos; below: embryos removed from the gametophytes
Here’s where it gets really interesting. Recall that one of the main functions of the female gametophyte, besides making the eggs, is to nourish the embryo. In other words, once the embryo starts to grow, it basically eats the gametophyte. It does this with the help of the suspensor, a column of disposable embryonic cells that push the main part of the new plant forward, into the gametophyte, so that it can absorb its nutrients. Once the embryo has established a distinct leafy end and root end, the root starts to grow back towards the suspensor and it crushes it. You can usually find the stringy dried up suspensor in the mature pine nut.
One of the things that makes pine nut embryos so adorable is the set of tiny needle leaves at their tips. When the embryo becomes a seedling, these will emerge to photosynthesize and take over the job of feeding the young plant.
Pine nuts at Christmas
Italian stone pines (Pinus pinea) are native to the European side of the Mediterranean coast. In Italy they occur on the northern half of both sides of the peninsula and in the heel of the boot. The range continues westward along the southern coast of France and into Spain and Portugal where native stands are scattered throughout the interior (Viñas et al., 2016). Pine nuts were never domesticated and are generally not even cultivated in orchards. They are usually harvested from natural stands, as they have been for tens of thousands of years in Southern Europe. There is even evidence from a Spanish cave that Neanderthals collected and presumably ate P. pinea seeds (Finlayson et al., 2006). Modern humans kept up the practice and many traditional foods from the region feature pine nuts.
Pine nuts have a distinctive conifer flavor dominated by pinene, limonene, hexanal, camphene, and careen (McGee, 2020), and they work well in both savory and sweet dishes where they hold their own against strong herbs and spices. There is of course pesto from Genoa in the heart of pine nut country, but also Italian cakes (pinolata) and Christmas cookies (pignoli). A specialty in parts of Provence is the sweet tarte aux pignons . In Catalonia, All Saints Day (November 1) is celebrated with pine nut confections called panellets. None of these traditional recipes includes chocolate — likely because they predate its arrival into Europe — but I really like to bake with a combination of chocolate, orange, and pine nuts, especially at Christmas.
Puff pastry tart with leeks, bleu cheese, arugula, and pine nuts
Investing in pine nuts
For a pine tree, the substantial energy allocated to female gametophytes is an investment in potential offspring with no guarantee of success. For us, it can be a substantial financial investment that may be increasingly costly for people and the planet as well. Pine nuts have always been more expensive than peanuts or almonds, but their price jumped this year for a variety of reasons (ProduceReport). Most pine nuts for sale in the United States come from stands of Korean pine growing in China. There, as everywhere, pine nut processing is unusually labor intensive and even dangerous, as rough heavy cones must be harvested by hand by skilled pickers who can navigate among the branches high above the ground. Seeds are then separated from the awkwardly knobby cones and the seed coats are removed from the female gametophytes. Pandemic-related safety measures and labor shortages have limited production, and the supply chain has been throttled, driving prices even higher. Meanwhile, a warming climate and a damaging insect pest have reduced yields (El Khoury et al., 2021). I’ll confess that I balked at the cost and used local pistachios in place of pine nuts in much of my baking this year.
The more I read about pine nut production the more concerned I became about worker protections and whether pine nut harvesting in natural stands could be sustained in the face of rising global demand. A conservation biologist working in Korean pine forests in Russia has written movingly about these highly diverse and fragile ecosystems, home to rare Amur tigers and other animals, and called for protections (Slaght, 2015).
Since a few western North American species produce large edible seeds, I looked for local harvesters who intentionally support both human and ecological communities. There are at least a couple of them, but neither had any product to sell this year. The future doesn’t look good for these businesses either, given the west’s megadrought and competition from lower-cost Chinese producers. Theirs is an investment against the odds and in favor of conserving an important cultural and ecological heritage.
The message of the pine nut
Besides their piney flavor and rich texture, what can we take from the precious little naked gametophytes that are now on my list of holiday plants? What message do I send to friends and colleagues along with my chocolate orange pine nut cake?
Pines have been around for about 150 million years, and conifers for twice that long (Rothwell et al., 2012, Jin et al., 2021), so their reproductive strategy can’t be that foolish. Their lineage persisted even when an asteroid slammed into our planet, causing the fifth mass extinction. If they don’t survive the Anthropocene, it won’t be because of their sex life. If anything, we should take their lesson to heart now more than ever. We can’t afford to wait until the last minute, like angiosperms do, to invest in future generations. It is time — past time, actually — to muster the courage and the will to dedicate all the resources we can to the preservation of the planet and our place in it. Otherwise, what hope do we have?
*note: Female gametophytes are more accurately called megagametophytes, and they derive from megaspores produced in megasporangia. Male gametophytes are really microgametophytes, pollen grains are microspores, and they are shed from microsporangia. In flowering plants, microsporangia are inside the anthers.
References and further reading
von Arnold, S., Clapham, D., & Abrahamsson, M. (2019). Embryology in conifers. Advances in Botanical Research, 89, 157-184.
El Khoury, Y., Noujeim, E., Bubici, G., Tarasco, E., Al Khoury, C., & Nemer, N. (2021). Potential Factors behind the Decline of Pinuspinea Nut Production in Mediterranean Pine Forests. Forests, 12(9), 1167.
Finlayson, C., Pacheco, F. G., Rodríguez-Vidal, J., Fa, D. A., López, J. M. G., Pérez, A. S., … & Sakamoto, T. (2006). Late survival of Neanderthals at the southernmost extreme of Europe. Nature, 443(7113), 850-853
Jin, W. T., Gernandt, D. S., Wehenkel, C., Xia, X. M., Wei, X. X., & Wang, X. Q. (2021). Phylogenomic and ecological analyses reveal the spatiotemporal evolution of global pines. Proceedings of the National Academy of Sciences, 118(20).
McGee, H. (2020). Nose dive: A field guide to the world’s smells. New York, NY: Penguin Press.
Meade, L. E., Plackett, A. R., & Hilton, J. (2021). Reconstructing development of the earliest seed integuments raises a new hypothesis for the evolution of ancestral seed‐bearing structures. New Phytologist, 229(3), 1782-1794.
Rothwell, G. W., Mapes, G., Stockey, R. A., & Hilton, J. (2012). The seed cone Eathiestrobus gen. nov.: fossil evidence for a Jurassic origin of Pinaceae. American Journal of Botany, 99(4), 708-720.
Rudall, P. J. (2021). Evolution and patterning of the ovule in seed plants. Biological Reviews, 96(3), 943-960.
Viñas, R. A., Caudullo, G., Oliveira, S., & de Rigo, D. (2016). Pinus pinea in Europe: distribution, habitat, usage and threats. European Atlas of Forest Tree Species; European Commission: Brussels, Belgium, 204.
Regular readers may have noticed that I (Katherine) sometimes go on a rant the week before Thanksgiving. This tradition is probably nothing more than a small annual outburst of snarky impatience that has accumulated over a long academic quarter, but I prefer to pretend that I am clearing space in my heart for gratitude. In past years I’ve gotten worked up about cooked celery, green bean casserole, and, most righteously, pecan pie. This year, my target is pickled peaches.
The wintertime pantry is a study in vegetable dormancy. Our shelves brim with structures plants use to store their own provisions. Each embryonic plant in a seed—the next generation of oats, quinoa, dry beans, walnuts—rests in the concentrated nutritive tissue gifted to it by its parent. The starchy flesh within the impervious shell of a winter squash is alive, as are apples, hopeful vessels of seed dispersal. Maple and birch syrup are stored energy made liquid and bottled. And then there are the so-called “root vegetables.” The term covers a surprisingly anatomically varied set of nutrient storage structures, only some of which are actual roots. Our familiar root vegetables represent only a sliver of global plant species diversity but nonetheless include the majority of contrivances herbaceous plants use in order to live to sprout another season: taproots, hypocotyls, stem tubers, root tubers, corms, and rhizomes. Raiding your root cellar for the ingredients for a roasted root vegetable medley, then, provides a great opportunity to turn your dinner prep into a botany lab. All you need is a knife and cutting board.
Roasted stacks of sweet potato and parsnip, painted with sage butter and roasted. See Katherine’s sweet potato post for the recipe.
The case for tree thinking
First we need to consider the taxonomy of our candidate botanical subjects. Taxonomy is the scientific practice of grouping related organisms in hierarchies of similarity. We shove the continuous variation of living things into discrete boxes labeled species, genus, family, order, and so on. Carl Linnaeus started the taxonomic naming system two hundred years before Watson and Crick identified the double helix shape of deoxyribonucleic acid (DNA), marking the beginning of the genomics era. Modern practitioners bring many types of data to bear—geography, fossils, genetics, morphology—toward the twin goals of illuminating the pattern of plant species evolution and defining groups based on common ancestry.
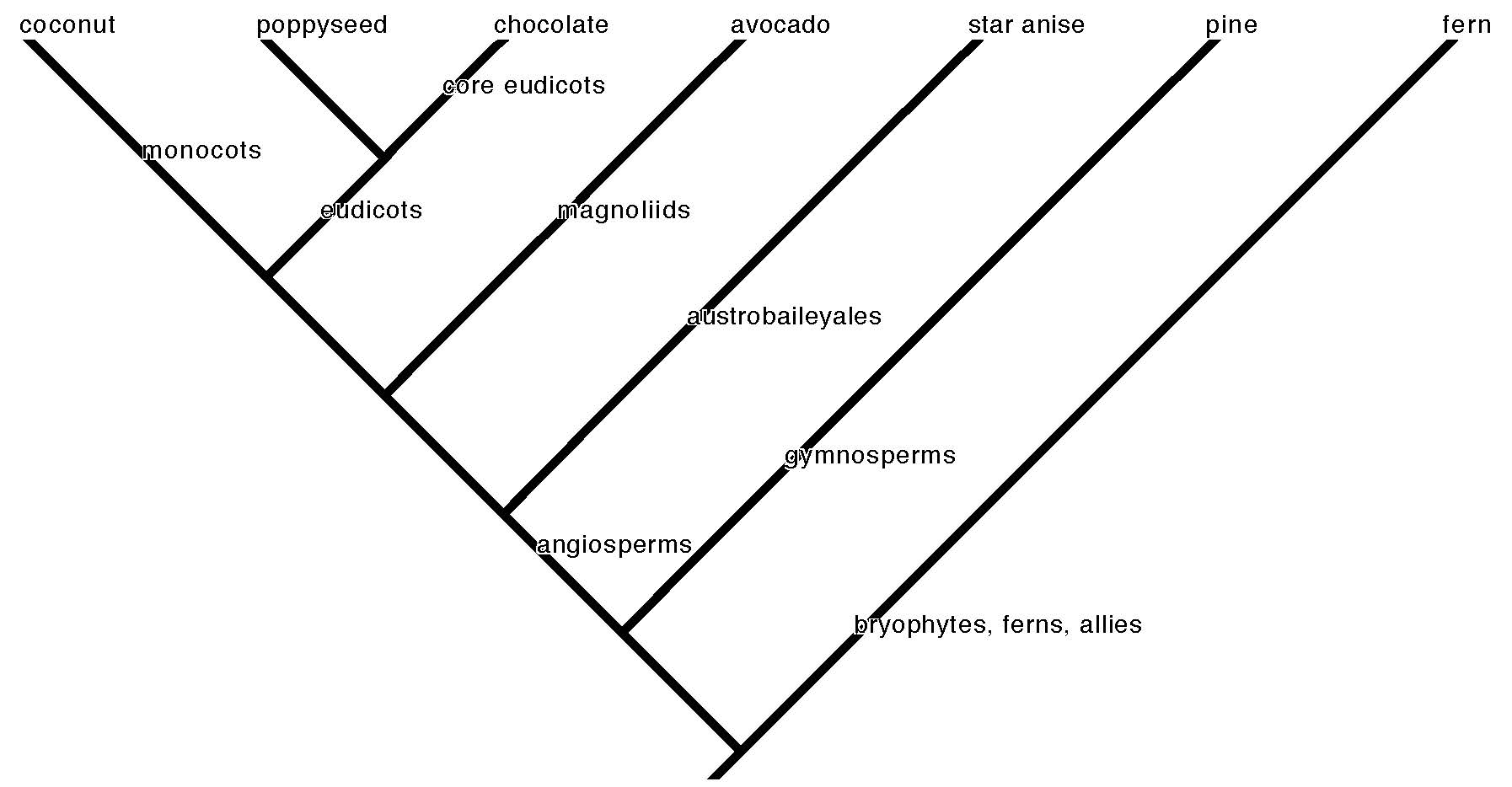
A phylogenetic tree of the major plant clades. Each branch point (node) represents the common ancestor of the organisms on the descendant branches. A single food plant species is shown here at the tip of each branch, a sort of mascot for its lineage.
The visual embodiment of this effort is the tree of life (cladogram) that represents the pattern of plant species evolution by common descent (phylogeny–see our primer on reading phylogenetic trees and using them to understand broad patterns in plant evolution). Each branching point on the tree is a node that represents a common ancestor of all the descendant taxa on the branches that come from it. The species are like the leaves on the tips of the branches. A schematic tree of life is the only illustration in Charles Darwin’s On the Origin of Species, the landmark book that provided the kernels of the core theories of evolutionary biology. Modern scientific convention tries to match old taxonomic names—because they are familiar and useful as a practical matter—with nodes on the tree of life. Small branches connect species to genus. Larger branches connect genera to families, families to order. The deep internal named nodes show the origin of the major clades. A clade is a group of organisms that descend from a common ancestor. A major clade is a significant branch on the plant tree of life that scientists have named for convenience of reference. You may remember some of these from biology class, like “monocot” and “dicot.” The former (monocots) has held up as a robust clade, but dicot is more complicated.
Green garlic, a monocot that stores its winter provisions as a bulb
As it happens, plant taxonomy before the advent of genetic data was reasonably accurate. Even though our understanding of plant species evolution is far from complete, genomic analysis has provided few big surprises about common ancestry of plant species and membership of taxonomic groups. Early taxonomists had the wisdom to rely primarily on similarity of reproductive structures—seeds, fruit, flowers, spores, cones—to circumscribe named groups. Reproductive structures tend to change more slowly over evolutionary time than do vegetative structures in plants. So one may expect to find a fair amount of coincident similarity among distantly related species in roots, shoots, and leaves.
This is where our categorization of root vegetables by taxonomy collides with our categorization of them by morphology. In short order we will organize our root vegetable species according to which structures the plant has chosen to amplify as a subterranean or soil-adjacent storage organ. This is not the same pattern as taxonomic organization. Grouping our root vegetables by taxonomy first helps us understand similarity and difference within and between groups of closely related plants—families, in this case. In doing so we can develop gestalt for the culinary qualities within plant families and appreciation for the evolution of plant diversity evident on our own dinner tables. Consider this intellectual nourishment, or perhaps the advent of a lens with which to view familiar foods anew.
Placing root vegetables on the plant tree of life
Around the globe humans utilize many dozens of plant species that bear underground (or near enough) storage structures. The most recent generations of people overwintering in the United States or Europe, however, chiefly engage with only a few. Perhaps only the most dedicated winter vegetable enthusiast will be familiar with all of the species on the following roster of root vegetables potentially available in Western grocery stores or farmer’s markets, although the list is unlikely to be exhaustive. I have organized the root vegetable species by families, and the families by major clade. Our list includes 15 of the 446 currently recognized plant families.
Whole sweet potatoes (Convolvulaceae)–NOT yams (Dioscoreaceae), NOT potatoes (Solanaceae), and NOT oca (Oxalidaceae)
Please take note of the disambiguation about the words “yam” and “potato.” The tubers marketed as “yams” in most American groceries are mostly actually sweet potatoes, which are also not potatoes. True yams are large tubers that are staples of tropical diets but relatively scarce in northern diets or groceries. In New Zealand the Andean oca is also known as “yam.” All of these are in different plant families.
Monocots:
Amaryllis family (Amaryllidaceae): onions and shallots (Allium cepa), garlic (Allium sativum), leeks (Allium ampeloprasum), and other alliums
Ginger family (Zingiberaceae): ginger (Zingiber officinale), turmeric (Cucurma longa)
Dioscoreaceae: true yams (several species in the genus Dioscorea), including the purple yam (ube; D. alata).
Sedge family (Cyperaceae): water chestnut (Eleocharis dulcis)
Arum family (Araceae): taro (Colocasia esculenta)
Eudicots: asterids
Goosefoot family (Amaranthaceae): beets (Beta vulgaris)
Carrots (taproot and leaves–which make a great pesto)
Now grab a carrot, parsnip, parsley root, salsify, or burdock root for your roast vegetable medley. These are the only true taproots on our list. The roots are much longer than they are wide and taper to a point. Thin lateral roots sprout from them in random locations or in discrete vertical lines. If you cut it open and examine it in cross section you see the tough core xylem (water conducting tissue) in the middle surrounded by a cortical layer (cambium) that separates the core from the sweet storage tissue (parenchyma) and sugar-moving phloem that surrounds it. Structurally supportive ray fibers radiate like spokes from the core. You have likely removed the aboveground greenery from these plants but should be able to tell or recall that it appears as if the leaves grow directly out of the crown of the taproots. They almost do. The anatomical stem on carrots and parsnips is a highly reduced disk on top of the taproot that serves as a bud-studded vascular transfer station, shuttling water and nutrients from the taproot into the leaves and flowering shoots.
Horseradish is also a taproot. A little bit grated into a sauce would make a delicious accompaniment to your roast vegetable medley. Incidentally, horseradish powder is the main ingredient in cheaper “wasabi” products available in American grocery stores, as the horseradish taste is superficially similar to that of true wasabi, which is also in the mustard family. Wasabi is also a root vegetable, but its underground storage structure is a rhizome, an underground stem, not a taproot.
A hypocotyl is a swollen fusion of taproot and stem base. The taproot portion is covered in fibrous secondary roots, most spectacularly in celery root. Leaf scars will be visible about these lateral roots, either surrounding the entire upper portions of the hypocotyl, as in celery root, or just at the top, as in beets and the mustard family hypocotyl vegetables (turnip, rutabaga, radish). All hypocotyl vegetables aside from beets are structurally straightforward but different from the taproots. A single layer of vascular tissue lays below the skin surface and penetrates into the storage tissue.
A rutabaga hypocotyl in the ground
Beets, however, are built from concentric rings of vascular tissue (xylem and phloem) and storage tissue (parenchyma), which is visible when the beet is cut in cross section. This ring structure is unique to the taxonomic order Caryopyllales, of which beets are a member. And as Katherine notes in her excellent beet post, the vibrant colors and earthy smell of beets are also unique. The former is due to betalain pigments, which are also unique to the Caryophyllales and distinct from the anthocyanin pigments present in all the other vegetables in our list (see our pigments post for a quick rundown of the most common pigments). The earthy smell is from a compound called geosmin. Beet is the only plant known to make it, and nobody knows why. Geosmin us usually produced by microbes in the soil and is liberated after rain to create that marvelous fresh smell after a storm.
chiogga beets show concentric vascular rings in dramatic fashion
Indidentally, our hypocotyl root vegetables here are all varieties, or subspecies, of species that also produce familiar leafy vegetables: rutabagas and the Russian or Siberian kales; turnips and Napa cabbages and mizuna; beets and Swiss chard; celery root and celery stalks or seeds. In each of these cases the variety produced for leaves has a much less pronounced hypocotyl than the variety produced as a root vegetable. Similarly, while the leaves on our hypocotyl root vegetables are all edible, they will be smaller and tougher than those on the varieties that have been bred for leaves.
Bulbs: onion, garlic, shallots, leek
red onion bulbs growing in a planter box
Onions, shallots, garlic and other alliums might be the most famous “root vegetables” of all, but their delicious parts are constructed entirely of swollen modified leaves. The papery tunicate covering surrounding the fleshy leaf bases are also constructed out of modified leaves, all arising from the basal plate (true compressed stem) that interfaces with the spindly roots on the bottom. The fleshy part of each garlic clove is a single fat modified leaf. Inside each garlic clove or onion bulb is an apical bud that will send up new leaves and flowering shoots. Everyone who has had onions and garlic sprout on them can observe this. You can of course plant these sprouting bulbs in the soil to make a new plant. A leek is a bit intermediate between a true bulb and a big herb. They call the lower white region of overlapping succulent leaf bases a “pseudobulb,” a nod to the messy continuous nature of biology and the difficulty with labels.
slices of leek pseudobulb, showing overlapping leaf bases
Unless you’re using a variety of “sweet” onion, which has been grown or bred to lack sulfurous aromatic compounds, you might tear up when you’re cutting onions and shallots. Cutting these bulbs volatilizes the irritating compounds that otherwise protect our favorite bulbs from pests.
Root tubers: sweet potatoes, cassava
A root tuber is an enlarged root that stores starch and other nutrients. Smaller lateral roots often branch from its surface and obtain water and soil nutrients. Raw sweet potatoes are readily available candidate root tuber ingredients for your botanical scrutiny and roast vegetable medley. Cassava is not, nor should it be, at least in root tuber form. Starch derived from cassava might be elsewhere in your pantry as tapioca.
A convenient aspect of our most commonly used root vegetables is that they require very little manipulation or preparation before they can be consumed. You don’t even have to peel your sweet potatoes before you cook them. Raw cassava tubers, however, are laced full to bursting with cyanide. They are the third most important source of calories throughout the tropics, behind corn and rice, but require extensive preparation before consumption to remove the cyanide, including grating, drying, leaching and cooking.
Cassava tubers develop underground from certain roots that become fleshy storage structures. They continue to acquire water and nutrients via smaller secondary roots that dot their surface. If the plant in question grows from a seed, then the harvestable storage root may develop from the taproot that grows from the seed. This, however, proves an inefficient way to farm these species, as many more storage roots can develop on a single plant when that plant is started from a shoot—a stem with leaves. This is where the visual heuristic of placing root vegetable species on the branches of the plant tree of life gets literal with sweet potatoes and cassava. The key factor is the presence of numerous nodes—leaves along the stem and their attendant axillary buds. Cassava and sweet potato are among the plant species that can generate roots from the buds in their leaf axils under the right conditions, namely being in contact with moist soil. Roots that develop from non-root tissue (like stems) are called adventitious roots. When several nodes of a shoot are planted in the soil, many adventitious roots will develop, of which some can become enlarged storage roots. In cassava the starting shoot is a cutting from a mature cassava plant. In sweet potatoes the starting shoot is called a slip. Slips grow from buds on the proximal (closest to the parent plant) end of sweet potato tubers. On sweet potatoes this is the end with the scar where the tuber was cut away from the parent plant.
A rhizome is a fleshy underground stem. It grows horizontally and sprouts new plants. Stems grow upward from buds near the soil surface, and roots grow from buds on the underside of the rhizome. It is structurally similar to stem tubers, like potatoes (see below), but it only grows horizontally, not in any direction, like a tuber. Rhubarb, asparagus, and irises also spread by rhizomes. If you decide to get out ginger or turmeric to flavor your vegetable medley, you’ll notice structural similarities to stem tubers, including nodes with buds.
Stem tubers: potato, sunchoke, jicama, yam
The eyes may or may not be the window into the soul, but they are our most conspicuous clue that potatoes are subterranean stem tubers, not roots. Katherine’s superb post on potato anatomy will walk you through this (potato) eye exam. Observe both ends of a potato. One end (the proximal end) bears the stump of the stolon (horizontal stem) that connected it to its mother plant. The other is tightly packed with small eyes that spiral out and around the potato. This is the growing (distal) end of the potato. New eyes originate at this end, so each eye is progressively older as you move toward the middle of the potato. Each eye contains a cluster of buds subtended by a semicircular leaf scar. The leaf in question was vestigial, translucent, and a remnant of it may still be present on your potato. Eyes are most easily visible on the “waxy” potato varieties (like Yukon Golds), which have less starch overall and a different ratio of types of starch than the “starchy” varieties (like Russets)–see Katherine’s post on potato starchiness for details.
potato eyes in spiral arrangement
The buds in each eye are axillary buds, structurally the same as Brussels sprouts. If your potato is exposed to enough light or warmth, the axillary buds will grow into new leafy stems, each of which can create a new potato plant. In this case your potato might also start synthesizing chlorophyll, turning it green. It will make toxic compounds at the same time, though, so if your potato is green you should either liberally peel it or wait to plant it in the spring.
Brussels sprouts on the stalk with residual leaf petioles. Brussels sprouts are spectacular axillary buds.
Nodes and buds are also easily visible on sunchokes, less so on jicama. True yams are actually structurally intermediate between rhizomes and stem tubers in that they might sprout adventitious roots. If you get your hands on an actual yam, instead of a sweet potato, you might see these.
sunchokes
Corms: taro, water chestnut (with a note on kohlrabi, which is not a corm)
A corm is yet another method by which plants have modified their stems to store starches and nutrients underground. The storage tissue is a swollen area of the stem above the roots and below the apical bud, from which leaves and flowers develop. Lateral buds on the stem produce modified leaves that produce a protective tunicate sheath around the starchy corm tissue. A thickened basal plate on the bottom interfaces with the roots and may sprout new corms (cormels). If you get canned water chestnuts or taro corms for your vegetable medley, these structures should be visible to you. Structurally, a taro corm is most similar to kohlrabi, which is what happened when plant breeders long ago took a weedy ancestral cabbage plant (Brassica oleracea) and bred for fat, bulbous stems. The leaf scars out the outside of a kohlrabi, and the nubbin of a root on the bottom, reveals that it is entirely stem.
Kohlrabi
The geophyte lifestyle
Potatoes are in the same genus (Solanum) as tomatoes (S. lycopersicum) and eggplants (S. melongena). The potato is the only one of these close relatives that hails from high in the Andes, where its underground tubers store the starches it needs to survive the harsh alpine conditions. This is a common ecological theme. Plants that create underground storage organs to withstand winter or seasons of drought are called geophytes. Even our short list of root vegetable species demonstrates that the geophyte lifestyle independently pops up all over the plant evolutionary tree, presumably in times and places where it may be adaptive. Even in just the Andes alone, potatoes are not the only domesticated geophyte crop with lowland relatives in the same genus devoid of starchy storage organs. Oca, confusingly called “yam” in New Zealand, where it was introduced in the mid-19th century, is otherwise known as Oxalis tuberosa. It makes stem tubers, like a potato. The specific epithet “tuberosa” separates it from non-geophyte species of Oxalis that are probably familiar to hikers and gardeners throughout the northern hemisphere. American health food stores sell dried maca hypocotyl (Lepidium meyenii) as a health food supplement, even though it is a staple crop throughout montane South America. Other Lepidium species are weedy little mustard plants. In the summer your garden may be teeming with flowering nasturtiums (Tropaeolum majus). You’ll notice a distinct lack of a fat, starchy stem tuber. Not so with mashua (Tropaeolum tuberosum).
branched taproot on a carrot
You should be well on your way at this point to getting your root vegetable medley into the oven. Finish peeling your vegetables, if you must, and dice them into approximately equally sized chunks. Toss them with a small amount of oil and salt. Add herbs if you would like. Spread them in a single layer on a baking tray or roasting pan and roast in the oven at 375 degrees Fahrenheit until they are tender, about 40 minutes. It is helpful to turn the pieces and move them around with a metal spatula halfway through the cooking time.
I like to serve these roast vegetables with some kind of sauce, often a strained yogurt mixed with salt and herbs. This is a dish filled by design with concentrated energy to maintain life through harsh seasons. The geophyte lifestyle is periodically useful for us all.