
Will seedless watermelons make us superhuman or turn our children into giants? Hardly, but they do give home cooks the power to count chromosomes without a microscope. Just a knife or a hard thunk on the sidewalk are enough to get a watermelon to spill its genetic guts.
If you were reading a Hearst Corporation newspaper in late 1937, you might have thought humanity would eventually be swallowed up by giant carnivorous plants, unwittingly unleashed by uncontrolled biotechnology. The San Francisco Examiner reported on November 21st of that year that the discovery of an “elixir of growth,” meant that “…science may at last have a grip on the steering wheel of evolution, and be able to produce at will almost any kind of species…” including “…a plague of man-eating ones.” In 1937 Americans had much more important things to worry about, just as we do now. Still, that discovery may in fact have threatened one cherished aspect of the American way of life by triggering the slow demise of late summer state fair watermelon seed spitting contests. It doubtlessly paved the way for seedless watermelons, and in 2014 the total harvest of seedless watermelons on American farms – nearly 700 thousand tons – outweighed the seeded watermelon harvest more than 13 to 1 (USDA National Watermelon Report). A similar pattern is emerging this year. Is there no stopping the attack of the seedless watermelons?

CLICK to read. Image from microfilm of an actual page in the San Francisco Examiner, published Sunday November 21, 1937. Found in the Media and Microtext Center of Stanford University Libraries.
And more important, how is it even possible to get seedless fruit from an annual plant? From a plant whose only mode of reproduction is through those very seeds? From a plant that cannot make suckers as bananas do and cannot be perpetuated endlessly through grafts like fruit trees and vines? Such is the challenge posed every single year by watermelons, but thanks to the “elixir of growth” discovered by Albert Blakeslee and subsequent work by Hitoshi Kihara, one of the most prominent agricultural geneticists of the 20th century, the world has an elegant solution. Breeders continually improve the varieties available, and consumer demand keeps growing, yet seedless watermelon production methods have remained essentially unchanged for three quarters of a century.

This watermelon definitely has seeds
Pollination without procreation
Seeds normally develop when pollen grains germinate on the stigma of a flower and pollen tubes grow down into the ovary to deliver sperm cells to the eggs inside the potential seeds (ovules). (see Jeanne’s fuller explanation here) The surrounding ovary is then stimulated to develop into a fruit. You might think that seeds could be avoided if pollination could be avoided. No pollen, no sperm, no seeds. But also no fruit. Large sweet fleshy fruits are expensive for a plant to produce, but the investment pays off if the fruit entices animals to disperse its seeds. Natural selection generally favors a high return on investment, and accordingly, watermelon fruits develop only if their flowers are amply pollinated and the chance of producing seeds seems high. (Although wild watermelon fruits are smaller than commercial varieties and not sweet, they still represent a serious investment of resources; see Paris 2015)
Preventing pollination is not a solution. It turns out, though, that a big load of pollen is enough to send the hormonal signals that promote watermelon fruit development, even if seeds ultimately never develop. What’s needed, then, is some way to trick the plant by providing a full pollen load while preventing the sperm in the pollen from fertilizing the egg and producing a viable embryo-filled seed.
Right now some of you are imagining thousands of microscopic condoms for the pollen tubes. It’s an amusing image, but the much more effective way to block embryo development is to start with defective egg cells. Plant egg cells are produced inside ovules (which would become seeds if fertilized) after a sequence of cell divisions that includes meiosis, the type of cell division that halves the number of chromosomes in a cell. Messing up meiosis will produce defective eggs. The geneticist Hitoshi Kihara knew that meiosis would be disrupted in every ovule of a plant if the plant body had three copies of each chromosome (a condition known as triploidy) instead of the usual two. On that point he seems to have been inspired by triploid seedless bananas (Crow, 1994). His great contribution was developing a technique to produce those triploid watermelon plants that would bear seedless fruit.

A seedless watermelon
Why triploids don’t make seeds
Humans normally have 23 pairs of homologous chromosomes, for a total of 46 chromosomes in most cells. One set of 23 is inherited from one parent, and the homologous set comes from the other. Similarly, watermelons normally have 11 pairs of chromosomes, for a total of 22. Having two copies of each chromosome makes us and watermelons diploid (Greek for 2-ply, written as 2N). Unless our parents are completely genetically identical (impossible in humans), the chromosomes in a pair are similar – “homologous” – but not identical. They carry the same sequence of genes, but will have different versions (alleles) of many of those genes. For example, both homologs of chromosome 6 in watermelon carry a gene influencing striping pattern, but a watermelon plant may have inherited the allele for defined stripes from one parent and the allele for diffuse stripes from the other.

A full set of human chromosomes, with homologous pairs brought together and digitally arranged in order of chromosome number.
When it comes time to make haploid gametes (eggs and sperm), the homologs have to be separated again so that each gamete carries a complete set of chromosomes, one of each pair. It’s almost funny how straightforward and logical the meiotic process is. The homologs find each other in the cell based on their similar gene sequences and become physically entwined. Each pair is then guided to the equator of the cell by tiny tubes (microtubules). When all the pairs are lined up properly, the homologs let go of each other and are pulled to opposite sides of the dividing cell. (A second round of division follows, but only the first round is critical for our watermelons.)
Like humans, watermelons are normally diploid; however (unlike humans) a plant with three copies of each chromosome (3N, or triploid) does perfectly well doing its everyday thing, growing and photosynthesizing. Plants in general are remarkably tolerant of extra chromosomes, and polyploidy is rampant in the plant world. For example, bread wheat is hexaploid (6N) and common commercial strawberries are octoploid (8N). The trouble starts with meiosis.

Small undeveloped seeds can be found in a seedless watermelon, but most people just eat these
Polyploids with even numbers of chromosomes, such as wheat and strawberries, can produce normal gametes (more on this below), but triploids (or pentaploids, etc.) mostly do not. When all three homologs are properly matched and lined up on the equator ready to separate, they cannot be divided equally. Sometimes only two homologs find each other, but then the third is left floating around on its own, which also prevents normal meiosis. With 11 sets of three homologs, in a given cell there may be a combination of indivisible triplets and pairs-plus-singles. In any case, the egg and its supporting cells fail to form, and the ovule remains small and soft and inoffensive enough to be ignored. The desired watermelon fruit develops in response to the pollen load, but the unwanted seeds never do.
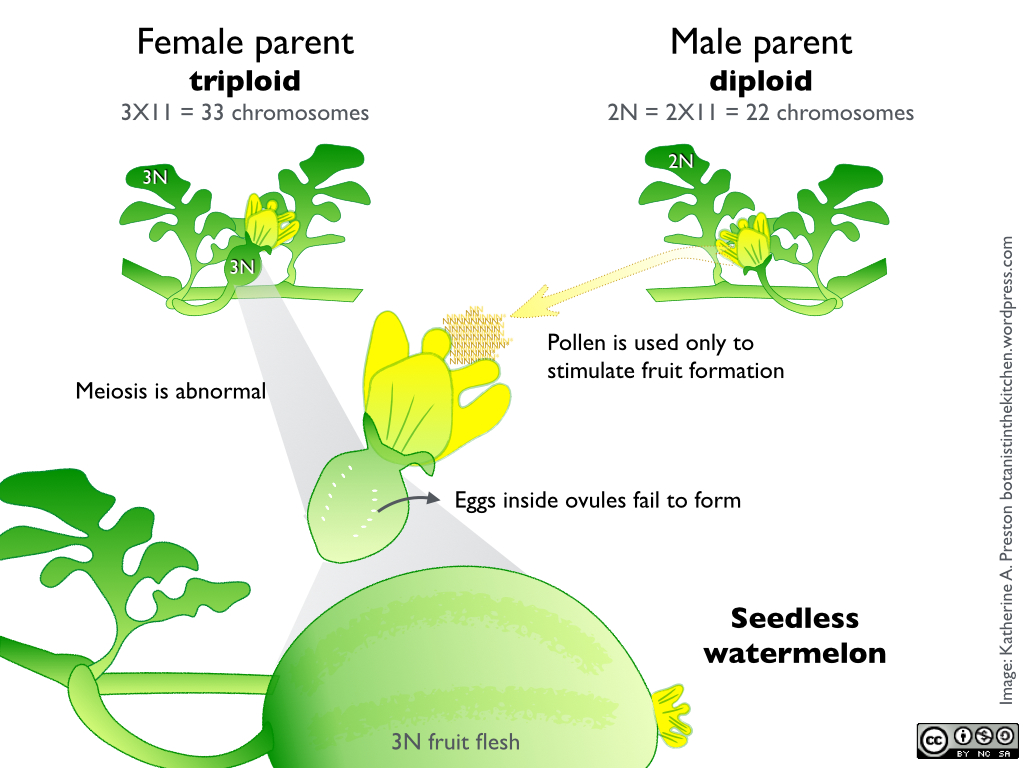
CLICK to enlarge. Plants that grow from triploid seeds grow normally, but they are sterile. Pollen from a diploid plant is used to stimulate fruit growth, but seeds never develop.
How we make triploids (but not giant rats)
Seedless triploids are beautiful in theory, but they don’t occur naturally in watermelons. How do breeders make a triploid plant from a diploid species? By bringing together a typical haploid (N) gamete from one parent and a diploid (2N) gamete from another. The most reliable way to produce diploid gametes is to start with a tetraploid parent (4N). Recall that even-numbered polyploids, unlike triploids, can make functional gametes through meiosis.
Tetraploid plants come about through various mechanisms, both natural and artificial. Sometimes ploidy is increased through hybridization between species (called “allopolyploidy,” as in bread wheat) and sometimes spontaneously within a species, even within a single plant (“autopolyploidy”).
The earliest written report of successful chemical induction of autopolyploidy was made in 1937 by Blakeslee and Avery, who were working at the Carnegie Institution at Cold Spring Harbor in New York. Following up on a tip from a colleague, they found that they could induce chromosome doubling by applying a plant-derived alkaloid called colchicine to either seeds or seedlings. Some of their experiments charmingly involved an atomizer “purchased in Woolworths for twenty cents.”

Applying colchicine with an atomizer from Woolworth. Excerpt from figure 5 of Blakeslee and Avery 1937.
What started innocently enough at Woolworths ended up splashed across the nation’s newspapers soon after Blakeslee and Avery’s discoveries became public. The reports were so overblown that the editor of the Journal of Heredity was moved to include a comment at the end of the published article to quell “over-enthusiastic popularizations” resulting from “the tidal wave of wierdly [sic] misleading publicity distributed by the Hearst newspapers” involving giant babies and wheat as tall as a pine tree. See below for an excerpt of his reassuring note. Yellow journalism aside, colchicine is toxic. Like many poisons, though, its properties are medically useful in low doses, and humans still take it for gout and some inflammatory diseases.
Colchicine works by interfering with mitosis, the kind of cell division that goes on constantly in our bodies, creating (ideally) identical cells for growth, maintenance, and healing. Before cells divide, they replicate their DNA so that each chromosome contains two copies of all the genetic material. As in meiosis, the chromosomes are moved to the middle of the cell by microtubules. In mitosis, however, the chromosomes line up single file and the two copies of the DNA are separated, not the homologs. A diploid cell becomes two diploid cells.
Colchicine-treated cells replicate their DNA as usual, but they are blocked from forming microtubules, so the cells do not divide; they just go on living with the extra copies of their chromosomes. What Blakeslee and Avery identified was the right amount of the chemical for the right length of time to prevent one round of cell division and induce tetraploidy but allow cells to start dividing again soon after that. With the correct protocol, plant parts that develop after colchicine treatment – including flowers – should all be tetraploid. Tetraploid flowers will produce diploid gametes (through meiosis) to be joined with haploid gametes from diploid plants to create triploid offspring.

CLICK to enlarge. Triploid watermelon seeds are made by crossing a tetraploid maternal plant with a diploid paternal plant. The resulting embryos are triploid. The fruits containing the seeds are tetraploid and not harvested for food. The triploid seeds are harvested for sale to growers.
It didn’t take long for Hitoshi Kihara to apply the new and exciting colchicine technique to watermelons, but it took a while to perfect it (Crow, 1994). One problem, as it turns out, is that tetraploid plants do still have some trouble making gametes, especially when all four copies of their chromosomes find each other and get tangled up. Breeders therefore select the most fertile tetraploid lines to develop, and over time meiosis becomes more normal. Once established, tetraploid lines can be maintained and propagated through seeds. There is no need to use colchicine again except to initiate a new tetraploid variety. Seed companies then use diploid plants to pollinate tetraploid flowers, which develop into fruit containing seeds with triploid embryos. We in turn buy those seeds to plant in our gardens as seedless varieties.
Growing triploids
With many very tasty seedless varieties available, why would anyone grow seeded watermelons these days? One reason is that triploid seeds cost about twice as much as diploid seeds because seed companies need to recoup their investment in developing tetraploid lines and generating seeds annually. Another consideration is the opportunity cost of growing seedless watermelon varieties. Alongside the triploid plants, farmers and gardeners have to plant diploid “pollinizer” plants, at a ratio of 2-to-1 triploids. Doing so ensures that there is enough pollen deposited on the stigmas of the triploid flowers to trigger fruit formation. Not only do farmers pay more for the triploid seeds, but they have to make room for the diploid vines to grow. Their extra cost is often passed along to consumers at the market or grocery store, who may be reluctant to pay the higher price.

CLICK to enlarge. From top left, moving clockwise, the stages of creating a seedless watermelon. To make a tetraploid line, colchicine is applied to diploid seedlings. Resulting tetraploid growth is allowed to flower and produce seeds. Those seeds are planted and the best versions are selected for several generations to establish a strong tetraploid line. Every year, seed producers fertilize tetraploid plants with pollen from diploid plants to create triploid seeds. Triploid seeds are sold to growers as seedless varieties. These plants must be pollinated by a diploid plant in order to make the fruit we eat.
Counting chromosomes and thinking of bees
Through the magic of colchicine, you have the power to tell a diploid from a triploid watermelon in your kitchen. If it has full-grown shiny black seeds, it’s a diploid. If it does not, it’s a triploid. That said, it is possible that in production, a tetraploid flower could be contaminated with pollen from another tetraploid flower and thus produce tetraploid seeds that accidentally get sold as triploids. However, according to the Cucurbit Breeding Project at North Carolina State University, tetraploids are usually bred to have a grey rind exactly so that any tetraploid seeds sneaking through would be evident to growers as soon as fruits started to develop.
I’ll admit that old fashioned seed-filled watermelons have their charms. Watermelon seeds are beautiful, especially against the deep red and sweet background of watermelon flesh. And the zone of seeds conveniently demarcates the heart. On the other hand, I love to contemplate the genetic gymnastics required to produce a seedless watermelon. If the biology of seedlessness is not enough to capture your own imagination, consider the bees. One study found that bees have to visit a triploid flower 16 to 24 times before it will produce a good fruit (Walters, 2005). That’s twice as many visits as a seeded variety requires. Humans may have escaped a nightmare plague of six-foot-long carnivorous caterpillars, but the bees really are in serious trouble, and we’d better do all we can to keep those bees happy and healthy and abundant.
Excerpt of a note by the editor of the Journal of Heredity, 1937, at the end of the article by Blakeslee and Avery:
“MANY readers of the JOURNAL will be interested, we feel sure, in some expansion and explanation of certain points not touched on in Dr. Blakeslee’s article, which have come up in connection with this most interesting subject. The tidal wave of wierdly misleading publicity distributed by the Hearst newspapers on November 21, has doubtless aroused hopes and fears that have no foundation whatever. Colchicine is not a “growth elixer,” and evidence is lacking as to its effects on animal chromosomes, so that doctors may still make their calls safe from attack by giant rats, and ladies are in no danger of being gobbled up by caterpillars suddenly gone carnivorous.
Colchicine is a narcotic alkaloid, related chemically to morphine and codeine. It is a very potent and very poisonous substance whose immediate effect on growing tissues even in very small concentrations is to produce stunting and distortion. It offers no Magical Royal Road to the production of new varieties of plants and animals. In spite of the cold water which must be thrown on over-enthusiastic popularizations, the effect which colchicine has on doubling the number of chromosomes in the cells makes it unquestionably one of the most important genetic discoveries of the year.”
References
Agricultural Research Service, USDA. (1971) Production of seedless watermelons. Technical Bulletin no. 1425. http://naldc.nal.usda.gov/download/CAT71326739/PDF
Blakeslee, A. F., & Avery, A. G. (1937). Methods of inducing doubling of chromosomes in plants by treatment with colchicine. Journal of Heredity, 28(12), 393-411. http://jhered.oxfordjournals.org/content/28/12/393.short
Crow, J.F. (1994) Hitoshi Kihara, Japan’s pioneer geneticist. Genetics 137: 891-894. http://www.genetics.org/content/137/4/891.full.pdf
Gama, R. et al. (2015) Microsatellite markers linked to the locus of the watermelon fruit stripe patternGenetics and Molecular Research 14 (1): 269-276 (2015) http://dx.doi.org/10.4238/2015.January.16.11
Paris, H. S. (2015). Origin and emergence of the sweet dessert watermelon, Citrullus lanatus. Annals of botany, mcv077.
Science opens the way to make our children giants. San Francisco Examiner (1937, Nov. 21). Media and Microtext Center, Stanford University Libraries.
USDA National Watermelon Report. Week ending Sept 4, 2015. http://www.ams.usda.gov/mnreports/fvdtvmelon.pdf
Walters, S. A. (2005). Honey bee pollination requirements for triploid watermelon. HortScience, 40(5), 1268-1270. http://hortsci.ashspublications.org/content/40/5/1268.short

Katherine,
Is it true that white “seeds” are unfertilized ovules? If so, how would they form given the fact that a fruit cannot develop unless a pollen grain triggers fruit development as you discussed above?
LikeLike
Errata: I meant “eggs,” not ovules.
LikeLike
Great question. First, egg cells develop as part of a larger structure called an ovule, which includes tissue from the mother plant and some cells resulting from meiosis. The outer layers of the ovule will develop to some degree before pollination occurs, and that’s what we see as the small white seeds. Further development of the seeds proceeds along with growth of an embryo inside, if eggs are normal and get fertilized. (Actually plants are pretty good at adjusting their allocation to reproduction at successive stages. They make more flowers than the number of fruits they can support, so they drop some. They can abort ovules or very young seeds if needed, and there is evidence that they preferentially support certain genotypes or high quality seeds.) Hope that helps.
LikeLike
Pingback: How giant pumpkins got so big: A Q&A with Jessica Savage | The Botanist in the Kitchen
Pingback: Botany Lab of the Month, Presidential Inauguration Edition: Saffron | The Botanist in the Kitchen
Pingback: Kiwifruit 1: Why are they so fuzzy? | The Botanist in the Kitchen
Please give me more about watermelon.
LikeLike
Sure thing! What do you want to know?
LikeLike
https://www.no-patents-on-seeds.org/index.php/de/patentfaelle/buschige-melone
https://register.epo.org/application?lng=de&number=EP13703094
LikeLike
Hello! I am curious how using a toxic substance such as
Colchicine would not harm the genetic makeup of the fruits or vegetables it is used to create? Is it removing nutrients or worse harmful to us in some way to eat them.
LikeLike
Thanks for your question, Beth. Colchicine treatment does not harm us at all since the fruits we eat don’t come from treated plants. Indeed, the treated plants are many generations in the past and located far away. Enlarge the diagram above to see the details. In short, the growing tip of the seedling is exposed to colchicine only long enough for cell division to be disrupted. Once those cells at the growing tip have twice the number of chromosomes as usual (are tetraploid), they will remain that way without any further colchicine treatment. The treated plant is allowed to flower, and its offspring for several generations are grown before being crossed with a diploid plant to create triploid seeds.
But you are right that working with colchicine is risky, and it is necessary to wear gloves and goggles and take other precautions to be sure it doesn’t get on your skin.
LikeLike